
Планктон, нектон, бентос | Биология. Реферат, доклад, сообщение, кратко, презентация, лекция, шпаргалка, конспект, ГДЗ, тест
Вода — среда обитания многих живых организмов. Немало организмов так и осталось жить в океане или в пресноводных водоёмах. Океан был и остаётся ареной развития бурной и многообразной жизни. Его населяет 300 тысяч видов животных и микроорганизмов и 30 тысяч видов растений.
Почему такая большая разница в видовом составе флоры и фауны Мирового океана? Дело в том, что животные заселяют всю толщу вод океана, а растения могут жить всего лишь до глубины 100 м, куда достигают солнечные лучи. И хотя по сравнению с толщей океана стометровая глубина кажется не очень значительной, именно этот слой обеспечивает жизнь в нём.
Живые организмы в океане и пресных водоёмах делятся на три группы: планктон, нектон и бентос.
Планктон (от греч. planktos — блуждающий) — это все организмы, населяющие пресные и солёные водоёмы и почти не способные к самостоятельному движению (рис.
Понятно, что представители фитопланктона — водоросли, способные к фотосинтезу, населяют океаны и моря лишь на глубине до 100 м. Фитопланктон служит пищей для зоопланктона и других водных животных.
| Рис. 167. Организмы, составляющие морской планктон: а—д — клетки водорослей; е — ночесветка; ж—и — мелкие ракообразные; к — медуза; л — гребневик; м — сагитта |
| Рис. 168. Горбатый кит |
Зоопланктон распространён по всей толще воды. Он представлен мелкими ракообразными, простейшими, кишечнополостными, икринками и личинками рыб. Для большинства планктонных организмов характерны приспособления для парения в толще воды: длинные выросты, газовые или жировые включения, студенистость тела и др.
Он представлен мелкими ракообразными, простейшими, кишечнополостными, икринками и личинками рыб. Для большинства планктонных организмов характерны приспособления для парения в толще воды: длинные выросты, газовые или жировые включения, студенистость тела и др.
Планктоном питаются различные мелкие рыбы (сардины, анчоусы и др.). И не только. Это излюбленное лакомство крупнейших морских животных — китов (рис. 168), которые поглощают громадное количество этих мельчайших обитателей океана, постоянно пропуская воду через пластины китового уса.
Нектон (от греч. nektos — плавающий) — активно плавающие водные животные, которые способны противостоять силе течения и перемещаться на значительные расстояния. К ним относятся 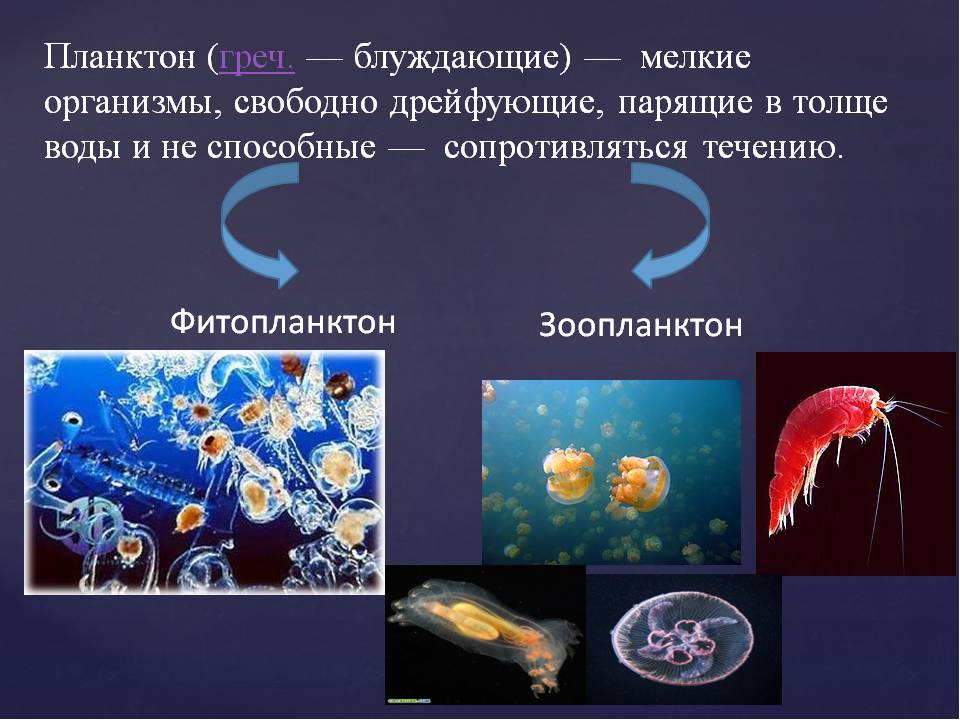 ru
ru
Бентос (от греч. benthos — глубина) — организмы, обитающие на грунте и в грунте дна рек, морей и океанов. К активно передвигающимся животным бентоса относятся морские звёзды, крабы, раки (рис. 169). Есть организмы, которые то всплывают, то лежат на дне, — камбалы (рис. 170) и скаты. Малоподвижны моллюски (гребешки, блюдечки). Ко дну прикрепляются устрицы и другие моллюски, а в грунт закапываются ланцетники. Основная масса бентоса живёт на мелководных участках морей.
| Рис. 169. Речной рак (а) и камчатский краб (б) |
| Рис. 170. Камбала |
Нектон организмы презентация
Скаты представители нектона
Нектон представители презентация
Информация о бентосе в океане
Планктон нектон бентос реферат краткое содержание
Для каких групп организмов вода является средой обитания?
Назовите известные вам группы океанических живых организмов, приведите примеры их представителей, опишите их особенности.

Что такое планктон, нектон и бентос? Приведите их примеры.

Организмы планктона
Планктон состоит из организмов, свободно живущих в толще воды и не способных противодействовать собственным движениям водной среды (течениям, конвекционным токам и т. п.) из-за отсутствия или относительно слабого развития у них органов движения. В систематическом отношении планктон делится на растительный планктон, или фитопланктон, и животный планктон, или зоопланктон.
В состав планктона входят, с одной стороны, голопланктические организмы, всю свою жизнь, включая также и период развития, проводящие вне связи с твердым субстратом, а с другой стороны — меропланктические организмы, которые определенный период своей жизни проводят на дне водоемов. К числу последних относятся, например, планктические личинки червей, иглокожих, моллюсков, ракообразных и других морских донных животных, отпочковывающиеся от полипов гидроидные медузы, а также многие живущие в прибрежной области организмы, цисты и покоящиеся яйца которых для дальнейшего развития опускаются на дно.
В зависимости от размера организмов планктон разделяется на следующие группы.
1. Ультрапланктон (бактерии)—величина организмов не превышает нескольких микронов, нижняя граница за пределами видимости.
2. Наннопланктон, или карликовый планктон (мельчайшие низшие растения и простейшие), — величина организмов измеряется микронами и десятками микронов; благодаря своим ничтожно малым размерам организмы наннопланктона проходят через самый густой шелковый газ, могут быть исследованы только при применении центрифугирования или камерного метода, поэтому эта группа организмов называется также центробежным, или камерным, планктоном.
3. Микропланктон (главная часть фитопланктона, а также инфузории, коловратки, мелкие ракообразные и т. п.)— величина организмов измеряется десятыми и сотыми долями миллиметра; улавливается планктическими сетями из густого шелкового газа или методом осаждения, поэтому называется также сетяным, или осадочным, планктоном.
4. Мезопланкон (крупные представители фитопланктона, главная часть зоопланктона морей) —величина организмов измеряется миллиметрами; улавливается планктическими сетями из редкого шелкового газа —сетяной планктон.
5. Макропланктон (высшие раки, медузы, пелагические черви и т. п.)—величина организмов измеряется сантиметрами, встречается исключительно в морях; улавливается большими планктическими сетями.
6. Мегалопланктон (многие сцифоидные медузы, крупные сифонофоры и т. п.) — величина организмов измеряется десятками сантиметров; встречается исключительно в морях.
Отличительная особенность планктических организмов — их способность находиться в воде во взвешенном состоянии — накладывает определенный отпечаток на их строение.
Еще интересные статьи по теме:
Личинки и куколки Polypedilum
Тело стройное, длиною 6—10 мм. Кольцевой орган расположен на нижней четверти первого ч
Brachionus plicatilis и Pedalia oxyure
Оба указанных вида коловраток обитают в планктоне малосоленых Балтийского и Азовского морей
Организмы бентоса
Бентос состоит из организмов, живущих на дне водоемов и не способных продолжительное время пПища водных животных
Для ознакомления с составом пищи производится вскрытие различных водных животных.
Черенки (Solen)
Черенки (Solen), относящиеся к морским двустворчатым моллюскам, живут в песке, в котором стр
Cercopagis
Виды Cercopagis, как и Podonevadne, распространены исключительно в Понто-арало-каспийской об
Планктон - пища морская, пища земная
В Вильфранш-сюр-мер, фешенебельном курорте Лазурного берега, работает одного из отделений Национального центра исследований Франции. Специалисты изучают морскую флору и фауну, и ведут иные океанографические исследования.
Одно из важнейших направлений – исследования океанического планктона.
Хотя вроде в рыбацкой сети улова не видно, на самом деле он – весьма обширен.
Просто речь идёт о мельчайших морских обитателях, которых практически не разглядеть невооружённым глазом.
Говорит Кристиан Сардэ, руководитель исследований:
“Это совсем крохи, размер не превышает полутора сантиметров, а еще есть сальпы, это свободноплавающие морские беспозвоночные, обитающие главным образом в поверхностных водах океана”.
Планктон – это пища для более крупных обитателей Мирового океана, любые колебания в его составе отражаются на всей экологической цепочке рано или поздно
Вот так планктон выглядит уже, когда на него смотрят “вооружённым взглядом”.
Всего морской планктон насчитывает более четверти миллиона видов живых существ: это креветки, мелкие рачки, водоросли и так далее. Ученые уточняют, что эти 250 тысяч – те, что изучены и описаны. А всего, по их подсчётам, планктон – это свыше миллиона таких существ.
Они относятся к разным ступеням эволюционной шкалы: у некоторых таких существ порой есть нервные окончания, сердце, кровеносная система.
“У каждого вида – своя, строго определённая роль. Вот этот мелкий рачок, к примеру, своего рода морское жвачное животное”, – замечает океанолог.
Упомянутые сальпы питаются фитопланктоном и сами служат пищей некоторым рыбам и морским черепахам.
Сальпы – это своего рода насос, который, прокачивая воду, помогает этому беспозвоночному, и двигаться, и добывать себе пищу.
Размножение сальпы сопровождается чередованием полового и бесполого поколений. В теле гермафродитной половой особи – бластозоида – из единственного яйца развивается особь бесполого поколения. По окончании развития оозооид покидает материнскую особь и приступает к почкованию новых бластозооидов
Сальпы – своего рода санитары морского леса, то есть водорослей, не давая последним размножаться в чрезмерных количествах.
Ещё одна особь, которая занимает учёных. Так называемый гребневик. Гребень – как плавник, он помогает сохранять нужную траекторию движения и не терять направления движения. Гребень может быть использован и как пропеллер, для ускорения передвижения в толще морской воды.
Планктон в той или иной форме обеспечивает потребности в пище почти всех обитателей Мирового океана, называется цифры 98 %.
“Более того, само наличие кислорода в атмосфере земли – это еще и наличие планктона. Он обеспечивает примерно 50 % кислорода, который содержится в земной воздушной смеси”, – резюмирует Кристиан Сардэ.
Планктон нуждается не только в изучении, но и бережном к себе отношении, подтверждают слова Кристиана Сардэ его коллеги.
Планктон впервые удалось изучить с маленькой яхты
Портал «Индикатор», 29.12.2020
Российские океанологи первыми в мире смогли изучить глубоководный планктон, плавая на небольшой яхте. Раньше подобные исследования проводились исключительно с больших научно-исследовательских кораблей, подчеркивается в пресс-релизе, поступившем в распоряжение редакции Indicator.Ru.
Около 95% всей обитаемой биосферы на Земле занимает глубоководная зона океана. Не зная, что там происходит, ученые не могут ни получить репрезентативную оценку экологического состояние планеты, ни решить многие научные и научно-прикладные задачи, такие как оценка ресурсов Мирового океана, моделирование потоков углерода и климатические модели. Главный обитатель глубин Мирового океана — планктон. Но изучение глубоководного планктона затруднено тем, что необходимые исследования требуют длительных экспедиций, значительных затрат времени и денег, а главное — опыта работ с глубоководными орудиями лова.
Эти проблемы сегодня очень сложно решить. Поэтому поддержаные грантами Российского научного фонда ученые из Лаборатории структуры и динамики планктонных сообществ Института океанологии им. П.П. Ширшова РАН решили работать принципиально новым способом — с использованием малых парусно-моторных кораблей с неограниченным районом плавания. Исследователи отправились в экспедицию на яхтенном научно-исследовательском судне в ноябре из Кейптауна (ЮАР). Закончили плавание они в декабре в Порт-Луи (Маврикий).
«Впервые в мировой практике нам удалось собрать глубоководный планктон до глубины 1500 метров не с большого научно-исследовательского судна, а с яхты длиной 20 метров. Ловы выполнялись самой большой планктонной сетью, которая эффективно ловит не только мезо-, но и макропланктон — планктонных рыб и креветок. Все 40 ловов на 14 станциях прошли без единого сбоя. Таким образом, была доказана принципиальная возможность проведения серьезных глубоководных исследований с малых судов. Эти суда не требуют долгого и дорогостоящего перегона к месту работ и обратно — они уже находятся в нужном районе», — рассказал руководитель проекта, заведующий лабораторией член-корреспондент РАН Александр Верещака.
Юго-западную часть Индийского океана, где работали российские океанологи, редко посещают ученые. В этот раз исследователям удалось сделать подробную съемку глубоководного планктона и собрать уникальный материал для изучения. Ученые обнаружили несколько новых видов и получили данные о запасах кормового планктона в этом районе океана, что важно для планирования промыслов. Все пробы с планктоном ученые фиксировали этиловым спиртом. Это позволит им применять самые современные методы генетических исследований, искать новые виды и роды, устанавливать связи местной фауны с фауной других районов океана. Кроме того, они подтвердили ранее полученные данные о кардинальной недооценке запасов глубоководных креветок. Их на 1–2 порядка больше, чем считалось ранее, а биомасса креветок на всех станциях сопоставима с биомассой всего остального планктона.
«Мы считаем, что успешное проведение экспедиции открывает новые стратегические перспективы изучения Мирового океана. Именно Мирового океана, а не отдельных небольших по меркам планеты районов, где сконцентрированы усилия океанологов в последнее время», — подчеркнул Верещака.
Живые организмы в океане — урок. География, 6 класс.
Представители флоры и фауны распространены не только на материках, но и в океане. Мировой океан является самой крупной средой обитания живых организмов.
На разнообразие видов живых организмов в океане оказывают влияние географическая широта и глубина. Количество видов животных и растений увеличивается от полюсов к экватору, а их масса, наоборот, уменьшается. В поверхностных водах Мирового океана обитает больше организмов, чем на огромной глубине. Это связано с большим количеством солнечного света и кислорода.
Растительность в океане значительно беднее, чем на материках. В Мировом океане очень мало цветковых растений. Водоросли — самая распространённая форма морской растительности.
Рис. \(1\). Морские водоросли
Фауна океанов богата и разнообразна. К самым крупным морским животным относятся млекопитающие (киты, дельфины, моржи, тюлени) и рыбы. Синий кит — крупнейшее животное Земли. В толще вод океана и на его дне обитают ракообразные, моллюски, морские звёзды, кораллы и многие другие.
Синий кит — крупнейшее животное Земли. В толще вод океана и на его дне обитают ракообразные, моллюски, морские звёзды, кораллы и многие другие.
Рис. \(2\). Морская звезда
Водные организмы разделяют на планктон, бентос и нектон.
Группа морских животных, которые обладают способностью противостоять силе течения и активно плавают в толще воды, называется нектоном (от греч. nektos — «плавающий», «плывущий»).
Рис. \(3\). Обыкновенный кальмар
Нектон — это киты, акулы, рыбы, морские змеи, кальмары, черепахи, моржи, тюлени, морские котики, пингвины и т. д.
Группа морских организмов, которые обитают на дне океана, называется бентосом (от греч. bentos — «глубина»).
Рис. \(4\). Коралл
К бентосу относятся губки, кораллы, морские черви, улитки, морские звёзды.
Группа мелких морских организмов, которые обитают в поверхностных водах и не способны сопротивляться течению, называется планктоном (от греч. planktos — «блуждающий»).
planktos — «блуждающий»).
Рис. \(5\). Бокоплав
Планктон — это микроскопические водоросли (фитопланктон), рачки, мальки рыб, личинки некоторых животных и ракообразных (зоопланктон), бактерии и простейшие. Планктон составляет \(70\) % от общей массы живого вещества Мирового океана.
Доклад: Планктон
Скачать на ЛитРес
Купить в Лабиринте
Планктон (от греч. plankt?s — блуждающий), совокупность организмов, населяющих толщу воды континентальных и морских водоёмов и не способных противостоять переносу течениями. В состав П. входят как растения — фитопланктон (в т. ч. бактериопланктон), так и животные — зоопланктон. П. противопоставляют населению дна — бентосу и активно плавающим животным — нектону. В отличие от последних, организмы П. не способны к самостоятельному движению или подвижность их ограничена. В пресных водах различают озёрный П.— лимнопланктон и речной — потамопланктон.
Растительные фотосинтезирующие планктонные организмы нуждаются в солнечном свете и населяют поверхностные воды, в основном до глубины 50—100 м. Бактерии и зоопланктон населяют всю толщу вод до максимальных глубин. Морской фитопланктон состоит в основном из диатомовых водорослей, перидиней и кокколитофорид; в пресных водах — из диатомовых, синезелёных и некоторых групп зелёных водорослей. В пресноводном зоопланктоне наиболее многочисленны веслоногие и ветвистоусые рачки и коловратки; в морском — доминируют ракообразные (главным образом веслоногие, а также мизиды, эвфаузиевые, креветки и др.), многочисленны простейшие (радиолярии, фораминиферы, инфузории тинтинниды), кишечнополостные (медузы, сифонофоры, гребневики), крылоногие моллюски, оболочники (аппендикулярии, сальпы, бочёночники, пиросомы), яйца и личинки рыб, личинки разных беспозвоночных, в том числе многих донных.
Размеры организмов П. колеблются от нескольких мкм до нескольких м. Поэтому обычно различают: наннопланктон (бактерии, наиболее мелкие одноклеточные водоросли), микропланктон (большинство водорослей, простейшие, коловратки, многие личинки), мезопланктон (веслоногие и ветвистоусые рачки и др. животные менее 1 см), макропланктон (многие мизиды, креветки, медузы и др. сравнительно крупные животные) и мегалопланктон, к которому относят немногих наиболее крупных планктонных животных (например, гребневик венерин пояс длиной до 1,5 м, медуза цианея диаметром до 2 м со щупальцами до 30 м, колонии пиросом длиной до 30 м и более 1 м в поперечнике и др.).
животные менее 1 см), макропланктон (многие мизиды, креветки, медузы и др. сравнительно крупные животные) и мегалопланктон, к которому относят немногих наиболее крупных планктонных животных (например, гребневик венерин пояс длиной до 1,5 м, медуза цианея диаметром до 2 м со щупальцами до 30 м, колонии пиросом длиной до 30 м и более 1 м в поперечнике и др.).
» Страница не найдена Гидробиологическое общество
Органом управления Гидробиологического Общества в период между съездами является Центральный совет Общества, в который входят (а) Президиум Центрального совета и (б) председатели региональных отделений Общества (подробнее см. Устав ГБО РАН).
ПРЕЗИДИУМ ЦЕНТРАЛЬНОГО СОВЕТА ГБО РАН
| Президент ГБО РАН: | |
| ГОЛУБКОВ Сергей Михайлович член-корреспондент РАН Зоологический институт РАН Университетская наб., 1 199034 Санкт-Петербург E-mail: Golubkov@zin.  ru ru | |
| Вице-президенты ГБО РАН: | |
| АДРИАНОВ Андрей Владимирович академик РАН Институт биологии моря ДВО РАН ул. Пальчевского, 17 690041 Владивосток | |
| ДГЕБУАДЗЕ Юрий Юлианович академик РАН Институт проблем экологии и эволюции им. А.Н. Северцова РАН Ленинский пр., 33 117071 Москва | |
| ПАВЛОВ Дмитрий Сергеевич академик РАН Институт проблем экологии и эволюции им. А.Н. Северцова РАН Ленинский пр., 33 117071 Москва | |
| Члены Президиума: | БОГАТОВ Виктор Всеволодович академик РАН Биолого-почвенный институт ДВО РАН пр. 100-летия Владивостока, 159 690022 Владивосток |
| ГЛАДЫШЕВ Михаил Иванович член-корреспондент РАН Институт биофизики СО РАН Академгородок 660036 Красноярск | |
| КОТОВ Алексей Алексеевич член-корреспондент РАН Институт проблем экологии и эволюции им.  А.Н. Северцова РАН А.Н. Северцова РАНЛенинский пр., 33 117071 Москва | |
| КРИКСУНОВ Евгений Аркадьевич член-корреспондент РАН Каф. Ихтиологии, Биологический факультет МГУ Воробьевы Горы 119899 Москва | |
| КРЫЛОВ Александр Витальевич д.б.н. Институт биологии внутренних вод РАН п/о Борок, Некоузский р-н 152742 Ярославская обл. | |
| МАТИШОВ Геннадий Григорьевич академик РАН Мурманский морской биологический институт РАН ул. Владимирская, 17 183010 Мурманск | |
| НАУМЕНКО Елена Николаевна д.б.н. АтлантНИРО ул. Дм. Донского, 5 236007 Калининград-обл. | |
| ТЕЛЕШ Ирина Викторовна д.б.н. Зоологический институт РАН Университетская наб., 1 199034 Санкт-Петербург | |
| Ученый секретарь ГБО РАН: | |
| Березина Надежда Александровна к.  б.н. б.н.Зоологический институт РАН Университетская наб., 1 199034 Санкт-Петербург E-mail: [email protected] | |
Члены Центрального совета ГБО РАН:Адрианов А.В.
Афанасьев Д.Ф.
Базова Н.В.
Безматерных Д.М.
Березина Н.А.
Богатов В.В.
Богданов В.Д.
Валькова С.А.
Гладышев М.И.
Голубков С.М.
Дгебуадзе Ю.Ю.
Думнич Н.В.
Зинченко Т.Д.
Ильмаст Н.В.
Итигилова М.Ц.
Карпинский М.Г.
Козлов О.В.
Копылов А.И.
Котов А.А.
Кренева К.В.
Крискунов Е.А.
Крылов А.В.
Лепская Е.В.
Лоскутова О.А.
Макаревич П.Р.
Малинина Ю.А.
Манушин И.Е.
Матишов Г.Г.
Мингазова Н.М.
Михайлова Л.В.
Науменко Е.Н.
Немова Н.Н.
Нечаев Д.Ю.
Павлов Д.С.
Пряничникова Е.Г.
Семерной В.П.
Соловых Г.Н.
Телеш И. В.
В.
Холмогорова Н.В.
Шадрин Н.В.
Шурганова Г.В.
Ядренкина Е.Н.
Яныгина Л.В.
Почетные члены ГБО РАН:- Богатов В.В. (Россия, Владивосток)
- Бульон В.В. (Россия, Санкт-Петербург)
- Васьковский В.Е. (Россия, Владивосток)
- Гулати Р.Д. (Нидерланды, Вагенинген)
- Дюмон Г. (Бельгия, Гент)
- Клековский Р.А. (Польша, Варшава)
- Ламперт У. (Германия, Плён)
- Морз Д. (США, Южная Каролина)
- Питч Т.У. (США, Сиэтл)
- Попченко В.И. (Россия, Тольятти)
- Протасов А.А. (Украина, Киев)
- Семенченко В.П. (Республика Беларусь, Минск)
- Телеш И.В. (Россия, Санкт-Петербург)
- Хлебович В.В. (Россия, Санкт-Петербург)
- Христофорова Н.К. (Россия, Владивосток)
- Шилова А.И. (Россия, Борок)
- Шуберт Г. (Германия, Росток)
- Шунтов В.П. (Россия, Владивосток)
- Эллиотт М. (Великобритания, Халл)
Йоланта Эйсмонт-Карабин, зав.
 гидробиологической станцией Экологического центра Польской академии наук, Миколайки, Польша.
гидробиологической станцией Экологического центра Польской академии наук, Миколайки, Польша.Пятьдесят пять лет назад в России было образовано Гидробиологическое общество. Сегодня эта научно-общественная организация (Гидробиологическое общество при Российской академии наук, или ГБО РАН) объединяет специалистов, осуществляющих исследования или занимающихся практической деятельностью по фундаментальным и прикладным направлениям гидробиологии и ихтиологии. В обществе состоят в основном научные сотрудники и инженерный персонал академических институтов, высших учебных заведений, рыбохозяйственных институтов, учреждений систем гидрометслужбы, санэпиднадзора, водного хозяйства и т.п. Главная цель общества — преодоление разобщенности специалистов, работающих в разных ведомствах и регионах России. Общество содействует контактам между гидробиологами России, стран СНГ и дальнего зарубежья. Кроме того, ГБО РАН способствует созданию условий для обмена информацией об исследованиях и достигнутых результатах, распространению и пропаганде наиболее перспективных методологических принципов и приемов выполнения исследований, анализу и интерпретации полученных научных материалов. Гидробиологическое общество объединяет усилия фундаментальных и прикладных направлений науки при решении важных задач, связанных с эксплуатацией водных ресурсов, и участвует в экспертизах по крупным водохозяйственным проектам с целью оценки экологических последствий их воплощения. Выполнение определенных уставом ГБО РАН функций осуществляется Президиумом и Центральным советом общества, региональными отделениями и в результате активной деятельности индивидуальных и коллективных членов общества.
Гидробиологическое общество объединяет усилия фундаментальных и прикладных направлений науки при решении важных задач, связанных с эксплуатацией водных ресурсов, и участвует в экспертизах по крупным водохозяйственным проектам с целью оценки экологических последствий их воплощения. Выполнение определенных уставом ГБО РАН функций осуществляется Президиумом и Центральным советом общества, региональными отделениями и в результате активной деятельности индивидуальных и коллективных членов общества.
Само Гидробиологическое общество непосредственно научные исследования не выполняет, но его центральные органы и, прежде всего, Президиум содействуют организации и проведению наиболее важных и перспективных работ. В решениях съездов общества определяются те приоритетные проблемы и направления исследований, на разработке которых целесообразно концентрировать внимание специалистов. Эти рекомендации служат для научных коллективов ориентирами и, как правило, находят отражение в конкретной гидробиологической тематике.
Однако, деятельность общества не сводится лишь к организационной работе его центральных органов и региональных отделений. В целом она является отражением активности всех отечественных гидробиологов, в том числе и не входящих в состав общества. В конечном итоге эта деятельность выражается в общих достижениях гидробиологии и заключается в развитии ее теоретических основ и разработке путей и методов решения актуальных вопросов, интересующих практиков. Поэтому целесообразно в краткой форме рассмотреть становление гидробиологических знаний, на фоне которого проявлялась деятельность Гидробиологическое общества, в свою очередь целеустремленно влиявшего на развитие науки.
Зарождение гидробиологической науки относится ко второй половине XIX века, но как фундаментальная наука экологического направления она складывалась на протяжении всего XX столетия, приняв современный вид преимущественно в его последней четверти. На отдельных этапах развития нашей страны эта наука претерпевала изменения, которые обусловливались как появлением и разработкой новых теоретических принципов, так и требованиями практики. Причем требования практики иногда приобретали достаточно жесткие формы. В определенной связи с развитием гидробиологии находились формы объединения специалистов в рамках «клуба по интересам», представленным в настоящее время Гидробиологическим обществом.
Причем требования практики иногда приобретали достаточно жесткие формы. В определенной связи с развитием гидробиологии находились формы объединения специалистов в рамках «клуба по интересам», представленным в настоящее время Гидробиологическим обществом.
Весь спектр гидробиологических исследований (по аналогии с большинством других наук) распадается на две ведущие ветви, одна из которых включает общие направления, вторая — прикладные вопросы [1]. Обе ветви находятся в тесном взаимодействии, причем первая разрабатывает теоретические основы, необходимые как для собственного развития, так и для эффективной деятельности второй. В то же время, исследования в рамках прикладной ветви не только обеспечивают решение разнообразных задач практики, но и стимулируют разработку новых общих (теоретических) проблем. На отдельных этапах истории гидробиологии лидирующее положение занимали общие или прикладные исследования, что было связано с повышением или ослаблением внимания как к гидробиологии в целом, так и к отдельным ее направлениям. Подобные изменения наглядно проявлялись в истории отечественной гидробиологии, особенно в 60-80-е годы прошедшего столетия, когда гидробиологический контроль качества водной среды сначала получил широкое развитие, а затем был существенно ограничен. Эти колебания отражались и на Гидробиологическом обществе.
Подобные изменения наглядно проявлялись в истории отечественной гидробиологии, особенно в 60-80-е годы прошедшего столетия, когда гидробиологический контроль качества водной среды сначала получил широкое развитие, а затем был существенно ограничен. Эти колебания отражались и на Гидробиологическом обществе.
При анализе развития общих направлений гидробиологических исследований в XX столетии выявляется интересная особенность, на которую в прошлом обращали мало внимания. Такие основные положения, характеризующие самостоятельную науку фундаментального ранга, как объект (предмет) исследования, цели и ведущий метод, не оставались постоянными, но существенно изменялись. В начале прошедшего столетия основное внимание уделялось качественным фаунистико-флористическим и биоценотическим исследованиям, что нашло отражение в классических монографиях С.А. Зернова [2] и К.М. Дерюгина [3]. В 20-е и 30-е годы XX столетия широкое применение получили количественные методы учета зообентоса и планктона, особенно в морских водоемах. Результаты исследований выражались в таких показателях, как число организмов и их биомасса, отнесенные к единицам площади водоема или объема воды. Поэтому объектами исследований в тот период считались либо организмы, биоценозы и типы водоемов [4], либо, по выражению Л.А. Зенкевича [5], водоемы со всей совокупностью жизненных явлений (включая биологическую продуктивность). Главными задачами, которые ставились перед гидробиологами, были изучение распределения организмов и биологическая продуктивность водоемов в том частном понимании, при котором исследователи ограничивались обсуждением показателей биомассы. Насколько широко такое представление о задачах исследований было распространено среди гидробиологов, видно из следующего примера. В связи с выходом первого тома трудов Всесоюзного гидробиологического общества группа ведущих гидробиологов выступила с программной статьей, в которой указывалось, что перед рассматриваемой наукой стоят три центральных проблемы: биологическая продуктивность водоемов, проблема биоценозов и проблема организма и среды [6].
Результаты исследований выражались в таких показателях, как число организмов и их биомасса, отнесенные к единицам площади водоема или объема воды. Поэтому объектами исследований в тот период считались либо организмы, биоценозы и типы водоемов [4], либо, по выражению Л.А. Зенкевича [5], водоемы со всей совокупностью жизненных явлений (включая биологическую продуктивность). Главными задачами, которые ставились перед гидробиологами, были изучение распределения организмов и биологическая продуктивность водоемов в том частном понимании, при котором исследователи ограничивались обсуждением показателей биомассы. Насколько широко такое представление о задачах исследований было распространено среди гидробиологов, видно из следующего примера. В связи с выходом первого тома трудов Всесоюзного гидробиологического общества группа ведущих гидробиологов выступила с программной статьей, в которой указывалось, что перед рассматриваемой наукой стоят три центральных проблемы: биологическая продуктивность водоемов, проблема биоценозов и проблема организма и среды [6].
Неудивительно, что в связи с отмеченными особенностями исследований, выполнявшихся на водоемах в первые десятилетия прошедшего столетия, проводившие их специалисты были еще тесно связаны с биологическими дисциплинами (в частности, зоологией, ботаникой и др.). Поэтому они обычно состояли в таких существовавших в то время научных объединениях, как Петербургское и Казанское общества естествоиспытателей, Московское общество испытателей природы и т.д. Однако расширение исследований, появление кафедр гидробиологии в вузах (в Сельскохозяйственной академии в Москве — 1914 г., Ленинградском и Московском университетах и др.) способствовали увеличению численности гидробиологов в академических и ведомственных научных учреждениях и иных организациях. Благодаря этому возникали объективные предпосылки для объединения гидробиологов в рамках специализированного общества.
Первым таким объединением явилось созданное в 1921 г. С.А. Зерновым в Москве «Общество исследователей воды и ее жизни». Как видно из самого названия, тематическая направленность деятельности общества целиком определялась теми представлениями, которые существовали в тот период по вопросу об объеме, целях и методе гидробиологических исследований. Печатным органом общества был «Русский гидробиологический журнал» (редактор А.Л. Бенинг), издававшийся Волжской биологической станцией в 1921-1930 г. в Саратове. Направленность работы журнала была такой же, как и общества, и совпадала с господствовавшими представлениями о гидробиологии как науке. В 1931 г. общество было преобразовано в секцию гидробиологии и ихтиологии в составе Московского общества испытателей природы. Благодаря этому положение с объединением специалистов в единую организацию «по интересам» внешне вернулось к состоянию, имевшему место до 1921 г. Но по существу ситуация резко изменилась. Дело в том, что в 20-е и 30-е годы XX столетия изучение водоемов в интересах развивающегося народного хозяйства значительно расширилось. На морях Баренцевом, Черном, Азовском, Каспийском, дальневосточных возникали рыбохозяйственные институты, ставшие крупными исследовательскими центрами.
Как видно из самого названия, тематическая направленность деятельности общества целиком определялась теми представлениями, которые существовали в тот период по вопросу об объеме, целях и методе гидробиологических исследований. Печатным органом общества был «Русский гидробиологический журнал» (редактор А.Л. Бенинг), издававшийся Волжской биологической станцией в 1921-1930 г. в Саратове. Направленность работы журнала была такой же, как и общества, и совпадала с господствовавшими представлениями о гидробиологии как науке. В 1931 г. общество было преобразовано в секцию гидробиологии и ихтиологии в составе Московского общества испытателей природы. Благодаря этому положение с объединением специалистов в единую организацию «по интересам» внешне вернулось к состоянию, имевшему место до 1921 г. Но по существу ситуация резко изменилась. Дело в том, что в 20-е и 30-е годы XX столетия изучение водоемов в интересах развивающегося народного хозяйства значительно расширилось. На морях Баренцевом, Черном, Азовском, Каспийском, дальневосточных возникали рыбохозяйственные институты, ставшие крупными исследовательскими центрами. Широкий размах приобрели работы на озерах и реках Европейской части страны и Сибири (Волга, Обь, Енисей, Лена, Колыма и др.), а также Украины, Казахстана, Средней Азии. Начинались исследования, связанные с назревавшим строительством крупных водохранилищ. В этой обстановке сужение рамок деятельности общественного объединения гидробиологов и ограничение их городом Москвой не способствовало преодолению разобщенности коллективов ученых. Однако созданию полноценного общества в тот период мешали, прежде всего, субъективные причины. Высшее руководство страны настороженно относилось к различным общественным объединениям, что создавало трудности по их организации.
Широкий размах приобрели работы на озерах и реках Европейской части страны и Сибири (Волга, Обь, Енисей, Лена, Колыма и др.), а также Украины, Казахстана, Средней Азии. Начинались исследования, связанные с назревавшим строительством крупных водохранилищ. В этой обстановке сужение рамок деятельности общественного объединения гидробиологов и ограничение их городом Москвой не способствовало преодолению разобщенности коллективов ученых. Однако созданию полноценного общества в тот период мешали, прежде всего, субъективные причины. Высшее руководство страны настороженно относилось к различным общественным объединениям, что создавало трудности по их организации.
| Академик С.А. Зернов — председатель «Общества исследователей воды и ее жизни» с 1921 по 1930 г. | Академик Б.Л. Исаченко — председатель ВГБО с 1947 по 1948 г. |
После Великой Отечественной войны обстановка несколько изменилась, и группа ведущих ученых Москвы, Ленинграда и Киева обратилась в Отделение биологических наук АН СССР с просьбой об организации Гидробиологического общества. Президиум АН СССР в мае 1946 г. возбудил ходатайство перед правительственными органами, и 2 апреля 1947 г. этот вопрос был решен положительно Постановлением Совета Министров СССР, в соответствии с существовавшей практикой. Указанную дату принято считать началом деятельности полноправного объединения гидробиологов страны в ее тогдашнем составе — Всесоюзного гидробиологического общества (ВГБО). Первыми председателями общества были акад. Б.Л. Исаченко (1947-1948 гг.) и акад. П.П. Ширшов (1948-1953 гг.), президентами состояли акад. Л.А. Зенкевич (1953-1970 гг.), чл.-корр. АН СССР Г.Г. Винберг (1970-1986 гг.), акад. АН СССР и Национальной академии наук Белоруссии Л.М. Сущеня (1986-1991 гг.). Начиная с 1991 г. президентом ВГБО, а затем — ГБО РАН (после распада СССР) избирается акад. А.Ф. Алимов.
Президиум АН СССР в мае 1946 г. возбудил ходатайство перед правительственными органами, и 2 апреля 1947 г. этот вопрос был решен положительно Постановлением Совета Министров СССР, в соответствии с существовавшей практикой. Указанную дату принято считать началом деятельности полноправного объединения гидробиологов страны в ее тогдашнем составе — Всесоюзного гидробиологического общества (ВГБО). Первыми председателями общества были акад. Б.Л. Исаченко (1947-1948 гг.) и акад. П.П. Ширшов (1948-1953 гг.), президентами состояли акад. Л.А. Зенкевич (1953-1970 гг.), чл.-корр. АН СССР Г.Г. Винберг (1970-1986 гг.), акад. АН СССР и Национальной академии наук Белоруссии Л.М. Сущеня (1986-1991 гг.). Начиная с 1991 г. президентом ВГБО, а затем — ГБО РАН (после распада СССР) избирается акад. А.Ф. Алимов.
| Академик П.П. Ширшов — председатель ВГБО с 1948 по 1953 г. | Академик Л.А. Зенкевич — президент ВГБО с 1953 по 1970 г. |
Сразу же после создания общества возникает его печатный орган — непериодический сборник «Труды ВГБО», первый номер которого вышел в 1949 г. Это издание существовало в течение сорока лет (до 1989 г.), всего было опубликовано 29 выпусков. С 1965 г. под эгидой Украинского отделения ВГБО в г. Киеве начал выходить «Гидробиологический журнал», оказавшийся после распада СССР за рубежами России. Помимо трудов и журнала, обществом и его отделениями издавались сборники научных материалов как через издательство «Наука», так и в регионах, а также методические руководства.
Это издание существовало в течение сорока лет (до 1989 г.), всего было опубликовано 29 выпусков. С 1965 г. под эгидой Украинского отделения ВГБО в г. Киеве начал выходить «Гидробиологический журнал», оказавшийся после распада СССР за рубежами России. Помимо трудов и журнала, обществом и его отделениями издавались сборники научных материалов как через издательство «Наука», так и в регионах, а также методические руководства.
| Член-корреспондент АН СССР Г.Г. Винберг — президент ВГБО с 1970 по 1986 г. | Академик Л.М. Сущеня — президент ВГБО с 1986 по 1991 г. |
| Академик А.Ф. Алимов — президент ГБО РАН с 1991 г. по 2014 г. | |
Возникновение общества сыграло существенную роль в развитии отечественной гидробиологии. Прежде всего, появление центральных органов общества в виде Президиума и Центрального совета стимулировало организацию региональных отделений, первыми из которых были Центральное (1947г.), Ленинградское (1948г. ), Киевское (1948г.), Латвийское (1949г.), Белорусское (1952г.) и др. К моменту созыва 1-го съезда ВГБО в 1965 г. в составе общества насчитывалось 27 региональных отделений, а в работе съезда приняло участие около 1000 человек из 94 городов бывшего СССР [7]. Такой интенсивный рост общества был связан, прежде всего, с расширением прикладных гидробиологических исследований, которые осуществлялись практически во всех окраинных и внутренних морях, в Мировом океане, во многих речных бассейнах, на озерах, водохранилищах, а также прудах. В связи с этим во второй половине 50-х гг. и в 60-е годы возникали или расширялись гидробиологические лаборатории или группы во многих академических и (особенно) ведомственных институтах, создавались специализированные кафедры в учебных заведениях.
), Киевское (1948г.), Латвийское (1949г.), Белорусское (1952г.) и др. К моменту созыва 1-го съезда ВГБО в 1965 г. в составе общества насчитывалось 27 региональных отделений, а в работе съезда приняло участие около 1000 человек из 94 городов бывшего СССР [7]. Такой интенсивный рост общества был связан, прежде всего, с расширением прикладных гидробиологических исследований, которые осуществлялись практически во всех окраинных и внутренних морях, в Мировом океане, во многих речных бассейнах, на озерах, водохранилищах, а также прудах. В связи с этим во второй половине 50-х гг. и в 60-е годы возникали или расширялись гидробиологические лаборатории или группы во многих академических и (особенно) ведомственных институтах, создавались специализированные кафедры в учебных заведениях.
Прикладные исследования осуществлялись по всему спектру гидробиологической проблематики, но основное внимание уделялось рыбохозяйственным и санитарно-водохозяйственным вопросам, изучению режима многочисленных новых водохранилищ, строившихся на главных реках страны, оценке экологических последствий осуществлявшихся и намечаемых проектов межбассейнового перераспределения водного стока и т. д. Среди рыбохозяйственных направлений большой объем исследований выполнялся по питанию и пищевым взаимоотношениям рыб и обеспеченности рыб кормовыми ресурсами, оценке рыбопродуктивности промысловых водоемов в связи с необходимостью увеличения промысла, повышению продуктивности озер, используемых для товарного рыбоводства, акклиматизации рыб и кормовых беспозвоночных, разработке теории продуктивности прудов. Расширялись работы по санитарно-гигиеническим и водохозяйственным вопросам, что обусловливалось необходимостью создания нормативной базы для контроля нарастающей антропогенной нагрузки на водные экосистемы со стороны промышленного и сельскохозяйственного производства и коммунальной сферы. В этой связи усиливались исследования водоемов питьевого назначения, биологического самоочищения водоемов, разрабатывались гидробиологические методы оценки качества водной среды и т.п. Невиданный размах приобрели начавшиеся еще в довоенные годы исследования гидробиологического режима крупных и крупнейших водохранилищ, возникавших в различных регионах страны: от рек Немана, Днестра и Днепра на западе до бассейнов рек Лены и Амура на востоке, а также в Казахстане, Средней Азии, Закавказье.
д. Среди рыбохозяйственных направлений большой объем исследований выполнялся по питанию и пищевым взаимоотношениям рыб и обеспеченности рыб кормовыми ресурсами, оценке рыбопродуктивности промысловых водоемов в связи с необходимостью увеличения промысла, повышению продуктивности озер, используемых для товарного рыбоводства, акклиматизации рыб и кормовых беспозвоночных, разработке теории продуктивности прудов. Расширялись работы по санитарно-гигиеническим и водохозяйственным вопросам, что обусловливалось необходимостью создания нормативной базы для контроля нарастающей антропогенной нагрузки на водные экосистемы со стороны промышленного и сельскохозяйственного производства и коммунальной сферы. В этой связи усиливались исследования водоемов питьевого назначения, биологического самоочищения водоемов, разрабатывались гидробиологические методы оценки качества водной среды и т.п. Невиданный размах приобрели начавшиеся еще в довоенные годы исследования гидробиологического режима крупных и крупнейших водохранилищ, возникавших в различных регионах страны: от рек Немана, Днестра и Днепра на западе до бассейнов рек Лены и Амура на востоке, а также в Казахстане, Средней Азии, Закавказье. В результате этих работ был накоплен большой объем оригинальных научных данных по этим техногенным водоемам, которые нуждаются в специальном анализе и обобщении.
В результате этих работ был накоплен большой объем оригинальных научных данных по этим техногенным водоемам, которые нуждаются в специальном анализе и обобщении.
Но эти перечисленные и другие прикладные направления гидробиологических исследований не могли бы выполняться с достаточной эффективностью при той теоретической базе, которая сложилась в науке к началу 50-х годов XX столетия. Господствовавший фаунистико(флористико)-биоценотический подход и описательные оценки продуктивности водоемов и биопродукционных процессов в них на основе учета биомассы организмов не позволяли удовлетворительно решать запросы практики при расчетах рыбной продукции, потребления кормовых ресурсов и обеспеченности ими популяций промысловых рыб, а также при прогнозных разработках количественных показателей состояния водных экосистем. Применение этого подхода приводило либо к далеким от реальности выводам, либо к прямым ошибкам (примеры см. в [8]). Благодаря активной деятельности ВГБО устаревшие представления об основах гидробиологии были в короткие сроки преодолены. В настоящее время основы гидробиологической науки опираются на результаты исследований, проведенных Г.Г. Винбергом (президент ВГБО в 1970-1986 гг.) и его научной школой, по определению пищевых потребностей, скоростей роста и энергетического обмена у гидробионтов. Эти представления о продуктивности водоемов позволяют количественно выражать продукционные процессы в водных экосистемах на всех трофических уровнях в виде потока энергии и «биотического баланса» [9]. Особенно широкий размах подобные исследования получили в России при выполнении работ по Международной биологической программе (МБП, 1964-1974 гг.). В рамках МБП, гидробиологические исследования осуществлялись по согласованным программам на разных типах водоемов (озера, водохранилища, пруды) в разных географических зонах: от Заполярья (Кольский полуостров) до Средней Азии и Закавказья, и от озер Литвы и Белоруссии до Восточной Сибири (Забайкалье) и Камчатки. Усилиями многих ученых был сделан ряд важных обобщений. Были выражены в виде уравнений функциональные зависимости скорости фотосинтеза планктона от содержания хлорофилла, зависимость его от среднегодовой концентрации фофора и прозрачности воды.
В настоящее время основы гидробиологической науки опираются на результаты исследований, проведенных Г.Г. Винбергом (президент ВГБО в 1970-1986 гг.) и его научной школой, по определению пищевых потребностей, скоростей роста и энергетического обмена у гидробионтов. Эти представления о продуктивности водоемов позволяют количественно выражать продукционные процессы в водных экосистемах на всех трофических уровнях в виде потока энергии и «биотического баланса» [9]. Особенно широкий размах подобные исследования получили в России при выполнении работ по Международной биологической программе (МБП, 1964-1974 гг.). В рамках МБП, гидробиологические исследования осуществлялись по согласованным программам на разных типах водоемов (озера, водохранилища, пруды) в разных географических зонах: от Заполярья (Кольский полуостров) до Средней Азии и Закавказья, и от озер Литвы и Белоруссии до Восточной Сибири (Забайкалье) и Камчатки. Усилиями многих ученых был сделан ряд важных обобщений. Были выражены в виде уравнений функциональные зависимости скорости фотосинтеза планктона от содержания хлорофилла, зависимость его от среднегодовой концентрации фофора и прозрачности воды. Показано, что эффективность продукционных процессов в водоемах в целом отражает соотношение между суммарными за год величинами продукции планктона и деструкции: в олиготрофных озерах баланс органических веществ часто отрицательный, а в эвтрофных и мезотрофных — близок к единице. Продукция автотрофов утилизируется в экосистеме в результате превращений ее в трофических цепях гетеротрофными организмами. Было установлено, что суммарная биомасса водных организмов в экосистемах различных водоемов возрастает пропорционально первичной продукции. Анализ биотических балансов, рассчитанных для разных по типу и географическому положению водоемов, позволил выявить ряд функциональных зависимостей, установить и количественно выразить генеральную схему потоков энергии в экосистемах. Особое внимание уделялось эколого-физиологическим исследованиям гидробионтов. Были определены количественные закономерности роста, размножения, обмена, питания животных и влияние факторов среды на эти процессы, решены многие сложные вопросы популяционной экологии гидробионтов, выявлены общие закономерности функционирования их сообществ.
Показано, что эффективность продукционных процессов в водоемах в целом отражает соотношение между суммарными за год величинами продукции планктона и деструкции: в олиготрофных озерах баланс органических веществ часто отрицательный, а в эвтрофных и мезотрофных — близок к единице. Продукция автотрофов утилизируется в экосистеме в результате превращений ее в трофических цепях гетеротрофными организмами. Было установлено, что суммарная биомасса водных организмов в экосистемах различных водоемов возрастает пропорционально первичной продукции. Анализ биотических балансов, рассчитанных для разных по типу и географическому положению водоемов, позволил выявить ряд функциональных зависимостей, установить и количественно выразить генеральную схему потоков энергии в экосистемах. Особое внимание уделялось эколого-физиологическим исследованиям гидробионтов. Были определены количественные закономерности роста, размножения, обмена, питания животных и влияние факторов среды на эти процессы, решены многие сложные вопросы популяционной экологии гидробионтов, выявлены общие закономерности функционирования их сообществ.
Использование теоретических положений продукционной гидробиологии позволило оценить продукционные особенности отдельных водоемов, представить направления возможных изменений их экосистем при загрязнении, ацидофикации или эвтрофировании, а также при других непреднамеренных или целенаправленных антропогенных воздействиях на водоемы.
Таким образом, в результате исследований отечественных гидробиологов — членов ВГБО и, прежде всего, школы Г.Г. Винберга, ядро которой находится в Зоологическом институте РАН, г. Санкт-Петербург, разработана теория биологической продуктивности водоемов и создано центральное направление гидробиологии — продукционная гидробиология, являющаяся в настоящее время основой этой науки [10].
В итоге, в гидробиологии фактически произошла смена ее составляющих: предмета (объекта), цели и основного метода исследования. Вместо качественных представлений о биоценозах и их фаунистико-флористическом составе и неопределенного понимания биологической продуктивности водоемов предметом исследования стали водные экосистемы, организуемые и регулируемые потоками вещества, энергии и информации, которые могут быть выражены в количественных показателях. Изменилось понимание задач (целей) гидробиологических исследований. По современным представлениям они заключаются не столько в описании органического мира водоемов, сколько в количественных оценках состояния водных экосистем и прогнозировании их возможных изменений под влиянием внешних (особенно антропогенных) факторов, а также в определении оптимальных условий и степени эксплуатации водоемов [11]. Новое содержание получили методы исследований. Хотя и прежде основным методом в гидробиологии считался количественный учет, но в настоящее время смысл этого термина существенно изменился. Место таких показателей, как численность и биомасса организмов, не дающих достаточно полного представления о количественной стороне протекающих в экосистемах процессов, заняли непосредственно определяемые и расчетные показатели: 1) продукция органического вещества за единицу времени, формирующаяся и преобразующаяся на разных трофических уровнях и выражаемая в универсальных энергетических единицах, 2) потоки информации, связывающие водную экосистему в единое целое и регулирующие ее динамику.
Изменилось понимание задач (целей) гидробиологических исследований. По современным представлениям они заключаются не столько в описании органического мира водоемов, сколько в количественных оценках состояния водных экосистем и прогнозировании их возможных изменений под влиянием внешних (особенно антропогенных) факторов, а также в определении оптимальных условий и степени эксплуатации водоемов [11]. Новое содержание получили методы исследований. Хотя и прежде основным методом в гидробиологии считался количественный учет, но в настоящее время смысл этого термина существенно изменился. Место таких показателей, как численность и биомасса организмов, не дающих достаточно полного представления о количественной стороне протекающих в экосистемах процессов, заняли непосредственно определяемые и расчетные показатели: 1) продукция органического вещества за единицу времени, формирующаяся и преобразующаяся на разных трофических уровнях и выражаемая в универсальных энергетических единицах, 2) потоки информации, связывающие водную экосистему в единое целое и регулирующие ее динамику.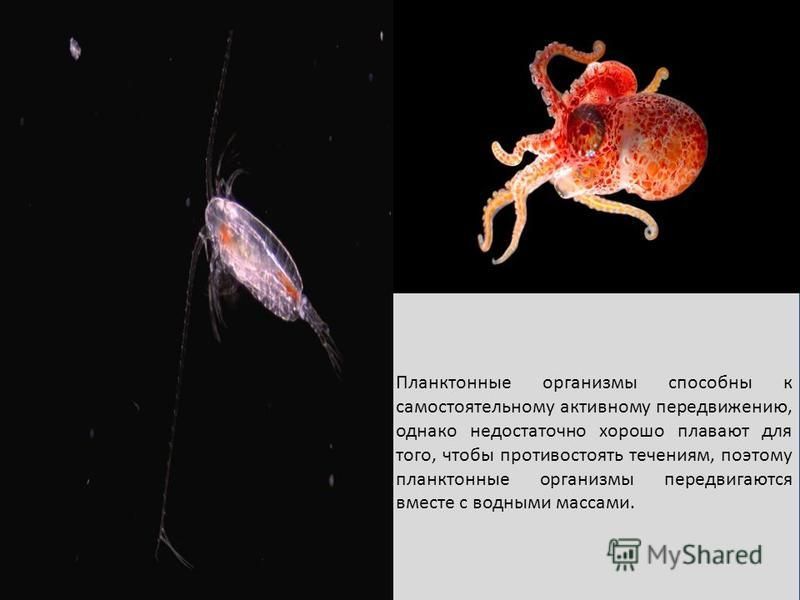 В комплексе динамических количественных методов видное место заняло моделирование функционирования водных экосистем.
В комплексе динамических количественных методов видное место заняло моделирование функционирования водных экосистем.
В связи с изменением предмета, цели и метода исследований гидробиология на современном этапе развития может быть определена как фундаментальная экологическая наука, осуществляющая изучение структуры и функционирования водных экосистем в их динамике, а также экологии организмов как составных элементов этих экосистем.
В течение последних 10-15 лет на теоретической базе продукционной гидробиологии была начата разработка важной для этой науки теории функционирования водных экосистем, основы которой были впервые изложены в статье А.Ф. Алимова [12] и детально развиты в его монографии [11]. Исследования в этом направлении приобретают все большую актуальность в связи с лавинообразным увеличением интенсивности эксплуатации водных ресурсов и практически повсеместным усилением антропогенного «давления» на водные экосистемы, нередко приводящим к их существенным преобразованиям, вплоть до полной деградации. Теория функционирования водных экосистем, вытекая из продукционного направления, выводит гидробиологические исследования на более высокий качественный уровень, придавая им новое концептуальное содержание. Она позволяет связать в единое целое протекающие в водных экосистемах потоки вещества и энергии с информационными потоками и тем самым придает понятию «водная экосистема» логическую и предметную завершенность. Поэтому дальнейшая разработка такой теории может рассматриваться как генеральное направление в развитии гидробиологии.
Теория функционирования водных экосистем, вытекая из продукционного направления, выводит гидробиологические исследования на более высокий качественный уровень, придавая им новое концептуальное содержание. Она позволяет связать в единое целое протекающие в водных экосистемах потоки вещества и энергии с информационными потоками и тем самым придает понятию «водная экосистема» логическую и предметную завершенность. Поэтому дальнейшая разработка такой теории может рассматриваться как генеральное направление в развитии гидробиологии.
Вместе с этими новыми тенденциями в общей проблематике гидробиологии происходят изменения и в прикладных направлениях этой науки. Помимо традиционных вопросов рыбохозяйственной и санитарно-гигиенической гидробиологии, все большее внимание исследователей и практических работников привлекают проблемы охраны окружающей среды. Решение этих проблем зависит от разработки и совершенствования гидробиологических методов слежения за экологическим состоянием водоемов (мониторинга) и методов его оценки, а также путей нейтрализации негативных антропогенных влияний (эвтрофикации, токсификации, ацидофикации и др. ) и восстановления (реабилитации) нарушенных водных экосистем. Можно ожидать, что в связи с нарастающим дефицитом пресной воды, ростом рекреационного использования водоемов и повышением их общего значения как важного элемента среды обитания человека, в ближайшей перспективе природоохранная проблематика станет доминирующей в прикладных гидробиологических исследованиях.
) и восстановления (реабилитации) нарушенных водных экосистем. Можно ожидать, что в связи с нарастающим дефицитом пресной воды, ростом рекреационного использования водоемов и повышением их общего значения как важного элемента среды обитания человека, в ближайшей перспективе природоохранная проблематика станет доминирующей в прикладных гидробиологических исследованиях.
Таким образом, в конце XX столетия особую актуальность в гидробиологии приобрели такие перспективные направления, как разработка теории функционирования водных экосистем и решение гидробиологических аспектов природоохранных проблем. Принимая во внимание значимость первого направления в теоретическом, а второго — в прикладном отношениях, можно полагать, что оба направления займут лидирующее положение в этой науке в первые десятилетия XXI века. В этой связи перед Гидробиологическим обществом встает задача по активному содействию в распространении и освоении названных новых представлений и направлений — подобно тому, как в недалеком прошлом оно способствовало распространению идей и методов продукционной гидробиологии.
Охарактеризованные выше принципиальные изменения в основах гидробиологии совпали по времени с коренными преобразованиями в стране. Перед российскими гидробиологами возникли сложные организационные задачи, которые приходилось решать в непростых условиях. Один из первых организационных шагов заключался в создании в 1993 г. Гидробиологического общества при Российской Академии наук (ГБО РАН), которое объединило российские отделения бывшего ВГБО, сохранив преемственность в работе. Несмотря на то, что вновь образованное общество оказалось в сложном финансовом положении из-за сокращения числа членов и общих экономических преобразований в стране, Президиуму и Центральному совету общества удалось сохранить эту научно-общественную организацию. Такие важные рычаги объединения исследователей, как организация и проведение совещаний, а также регулярный созыв съездов с сохранением прежней периодичности (раз в пять лет) и нумерации, сыграли главную роль в этом интеграционном процессе.
Проблемы теоретической и прикладной гидробиологии по мере ее становления и итоги выполнявшихся в этом направлении исследований широко обсуждались на многих совещаниях, созывавшихся Гидробиологическим обществом, и на всех его съездах. Очередной, 7-ой съезд ГБО РАН был проведен в 1996 г. в г. Казани, 8-ой съезд — в г. Светлогорск Калининградской области. В обоих случаях было издано по три тома материалов съездов. В 1995 г. обществу удалось в качестве одного из соучредителей организовать выпуск печатного органа — журнала «Биология внутренних вод» (издательство «Наука»). В настоящее время обсуждается вопрос о возрождении издания «Русского гидробиологического журнала» в электронной версии.
Очередной, 7-ой съезд ГБО РАН был проведен в 1996 г. в г. Казани, 8-ой съезд — в г. Светлогорск Калининградской области. В обоих случаях было издано по три тома материалов съездов. В 1995 г. обществу удалось в качестве одного из соучредителей организовать выпуск печатного органа — журнала «Биология внутренних вод» (издательство «Наука»). В настоящее время обсуждается вопрос о возрождении издания «Русского гидробиологического журнала» в электронной версии.
Прошедшие 7-ой и 8-ой съезды явились важными переломными вехами в деятельности ГБО. Это доказывают следующие цифры и факты. В работе 7-го съезда приняло участие 85 делегатов, представлявших 620 членов общества, и было заслушано 116 докладов. В то же время, на 8-ой съезд прибыло 285 участников, в том числе 233 делегата, представлявших более чем 800 членов общества, и было заслушано 260 докладов. На этом съезде в качестве гостей и докладчиков присутствовали гидробиологи из Белоруссии, Литвы, Польши Украины, что свидетельствует о возрождении международных связей общества. Таким образом, можно считать, что Гидробиологическому обществу удалось преодолеть кризисные дезинтеграционные явления в науке 90-х годов XX столетия, и оно вступило в XXI век обновленным как в теоретическом и прикладном отношениях, так и организационно.
Таким образом, можно считать, что Гидробиологическому обществу удалось преодолеть кризисные дезинтеграционные явления в науке 90-х годов XX столетия, и оно вступило в XXI век обновленным как в теоретическом и прикладном отношениях, так и организационно.
ЛИТЕРАТУРА1. Винберг Г.Г. Гидробиология как экологическая наука // Гидробиологический журнал. 1977. Том 13, № 5.
2. Зернов С.А. Об изучении жизни Черного моря // Записки АН, VIII серия, физико- математическое отделение. 1913. Том 32, № 1.
3. Дерюгин К.М. Фауна Кольского залива и условия ее существования // Записки АН, VIII серия, физико-математическое отделение. 1915. Том 34, № 1.
4. Зернов С.А. Общая гидробиология. Москва-Ленинград: Гос. изд-во биологической и медицинской литературы, 1934. (2-ое издание: 1949).
5. Зенкевич Л.А. Фауна и биологическая продуктивность моря. Том 1. Мировой океан. Москва: Советская наука, 1951.
6. От редакционного совета // Труды Всесоюзного гидробиологического общества. 1949. Том 1.
1949. Том 1.
7. Скрябина Е.С. Всесоюзное гидробиологическое общество. Справочник. Москва: Наука, 1984.
8. Кудерский Л.А. К вопросу о методике определения рыбной продуктивности водоемов // Известия Карельского и Кольского филиалов АН СССР. 1958. Вып. 5.
9. Алимов А.Ф. К пятидесятилетию Гидробиологического общества // Гидробиологические исследования в бассейне Атлантического океана. Том 1. Пресноводная гидробиология. Калининград: АтлантНИРО, 2000.
10. Алимов А.Ф. Введение в продукционную гидробиологию. Ленинград: Гидрометеоиздат, 1989.
11. Алимов А.Ф. Элементы теории функционирования водных экосистем. Санкт-Петербург: Наука, 2000.
12. Алимов А.Ф. Основные положения теории функционирования водных экосистем // Гидробиологический журнал. 1990. Том 26, № 6.
XI съезд Гидробиологического общества при РАН в СФУ
Объединенный Пленум Научного совета по гидробиологии и ихтиологии РАН, Гидробиологического общества при РАН и Межведомственной ихтиологической
(25 марта 2017 года)
С чем я плаваю? (Служба национальных парков США)
Штормовой пляж в Национальном парке АкадияNPS Фото
Вы когда-нибудь плавали и задавались вопросом, что может жить в этом пруду или озере? А океан?
В водах национального парка Акадия можно встретить рыб, ракообразных, амфибий и многих других живых существ. Но думали ли вы обо всем, что живет в воде, чего вы не можете видеть?
Но думали ли вы обо всем, что живет в воде, чего вы не можете видеть?
В каждом водоеме Национального парка Акадия живут бесчисленные микроскопические организмы, известные как планктон.Планктон — это микроскопические растения и животные, которые дрейфуют по воде, не плавая самостоятельно. В одной столовой ложке океанской воды могут быть сотни тысяч планктонных организмов. Существует два основных вида планктона – фитопланктон и зоопланктон.
Фитопланктон
Планктон, способный к фотосинтезу, называется фитопланктоном. Как и другие растения, они используют солнечный свет для создания энергии. Фитопланктон составляет основу морской пищевой сети, являясь источником пищи для других планктонов и более крупных животных.Фитопланктон также важен для количества кислорода, который они производят.
Зоопланктон
Планктон, который потребляет другие живые существа, чтобы выжить, известен как зоопланктон. Зоопланктон обычно поедается почти всеми личинками рыб, а это означает, что они являются важной частью пищевой сети, которая в конечном итоге ведет к нам, людям. Без планктона, который потребляют эти личинки рыб, вся экосистема, основанная на планктоне, не выживет.
Без планктона, который потребляют эти личинки рыб, вся экосистема, основанная на планктоне, не выживет.
Хотя планктон может быть очень важной частью водных экосистем Акадии, слишком много хорошего может быть плохим.При наличии слишком большого количества питательных веществ фитопланктон может выйти из-под контроля, что приведет к так называемому цветению водорослей. Эти цветы могут быть вредны для рыб, млекопитающих и даже людей.
Однако без цветения водорослей плавать с планктоном совершенно безопасно. Наслаждайтесь плаванием в озерах и прудах острова Маунт-Дезерт и в холодных чистых водах Атлантического океана, теперь зная, в чем вы плаваете.
Сообщество Планктона Вата | Plankton Wat
Сообщество Plankton Wat | Планктон Ват ← еще отThrill Jockey Records ••• показывай меньше
Каждую пятницу получайте свежие музыкальные рекомендации на свой почтовый ящик.
Подпишитесь на Plankton Wat, чтобы увидеть это сообщение Подпишитесь на Plankton Wat, чтобы увидеть это сообщение
Следовать Выучить большеПрикрепленная Почта
Новый прямой эфир от Эксклюзивное сообщение для подписчиков Plankton Wat только что запустил подписку В вашу коллекцию добавлен новый выпуск только для подписчиков Эксклюзивное сообщение для подписчиков Новый релиз добавлен в вашу коллекцию Новый релиз от Новый релиз от
Подпишитесь на Plankton Wat, чтобы увидеть это сообщение Подпишитесь на Plankton Wat, чтобы увидеть это сообщение
Следовать Выучить большеПрикрепленная Почта
Новый прямой эфир от Эксклюзивное сообщение для подписчиков Plankton Wat только что запустил подписку В вашу коллекцию добавлен новый выпуск только для подписчиков Эксклюзивное сообщение для подписчиков Новый релиз добавлен в вашу коллекцию Новый релиз от Новый релиз от
избранный трек:
Планктон Ват
Портленд, Орегон
Портлендский музыкант Дьюи Махуд — один из самых разносторонних и плодовитых музыкантов современного психоделического андеграунда.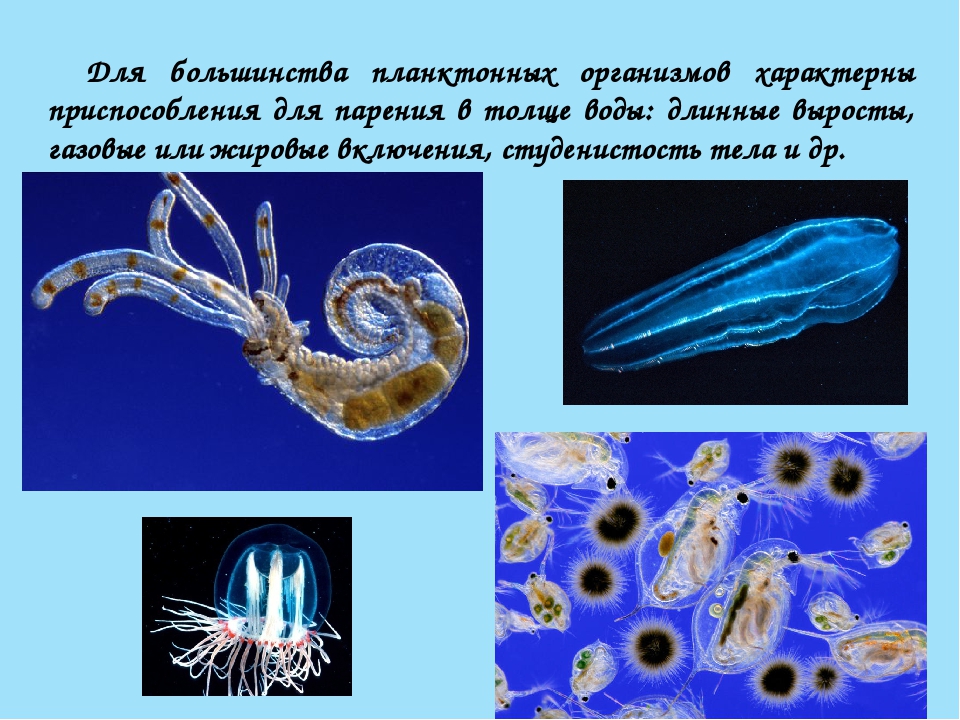 Как Планктон Уот, он исследовал дальние рубежи фолка и синтезированного дрифта с середины нулевых, и его преданность импровизации превыше всего, его неугомонный исследовательский дух заслужили ему место темной лошадки в континууме современного звукового искусства. исследователи.
Как Планктон Уот, он исследовал дальние рубежи фолка и синтезированного дрифта с середины нулевых, и его преданность импровизации превыше всего, его неугомонный исследовательский дух заслужили ему место темной лошадки в континууме современного звукового искусства. исследователи.
(PDF) Изменение климата и морской планктон
20 Greve, W.и другие. (2001) Прогнозирование сезонности
зоопланктона Северного моря. Senckenbergiana maritima 31, 263–268
21 Mackas, D.L. и другие. (1998) Междекадные изменения в сроках развития
популяций Neocalanus plumchrus на океанской станции P в
субарктической северной части Тихого океана. Может. Дж. Фиш. Аква. науч. 55, 1878–1893
Может. Дж. Фиш. Аква. науч. 55, 1878–1893
22 Гертен Д. и Адриан Р. (2000) Климатические изменения весной
Динамика планктона и чувствительность мелководных полимиктовых озер к
Североатлантическому колебанию.Лимнол. океаногр. 45, 1058–1066
23 Уиндер М. и Шиндлер Д.Е. (2004) Изменение климата расцепляет
трофических взаимодействий в водной экосистеме. Экология 85, 2100–2106
24 Эдвардс, М. и Ричардсон, А.Дж. (2004) Влияние климатических
изменений на фенологию планктонного сообщества и
трофических несоответствий. Nature 430, 881–884
25 Стенсет, Н. К. и Мистеруд, А. (2002) Климат, изменение фенологии,
и другие черты истории жизни: нелинейность и несоответствие
окружающей среде.проц. Натл. акад. науч. U. S. A. 99, 13379–13381
26 Cushing, D.H. (1972) Производственный цикл и численность морской
рыбы. Симп. Зоол. соц. Лонд. 29, 213–232
27 Шеффер, М. и Карпентер, С.Р. (2003) Катастрофические изменения режима в
экосистемах: связь теории с наблюдениями. Тенденции Экол. Эвол. 18,
Тенденции Экол. Эвол. 18,
648–656
28 Steele, J.H. (2004) Режимные сдвиги в океане: согласование наблюдений и теории. прог. океаногр.60, 135–141
29 Венрик, Э.Л. и другие. (1987) Климат и хлорофилл а: долгосрочные тенденции
в центральной части северной части Тихого океана. Science 238, 70–72
30 Beaugrand, G. (2004) Изменение режима Северного моря: доказательства, причины,
механизмы и последствия. прог. океаногр. 60, 245–262
31 Дуарте, К.М. и другие. (1992) Неопределенность обнаружения изменения моря. Природа
356, 190
32 МГЭИК (2001) Изменение климата 2001: Научная основа. Вклад
Рабочей группы I в Третий оценочный отчет Межправительственной группы экспертов по изменению климата
, Cambridge University Press
33 Caldeira, K.и Wickett, ME (2003) Антропогенный углерод и
рН океана. Nature 425, 365
34 Riebesell, U. (2004) Влияние обогащения CO
2
на морской фитопланк-
тонны.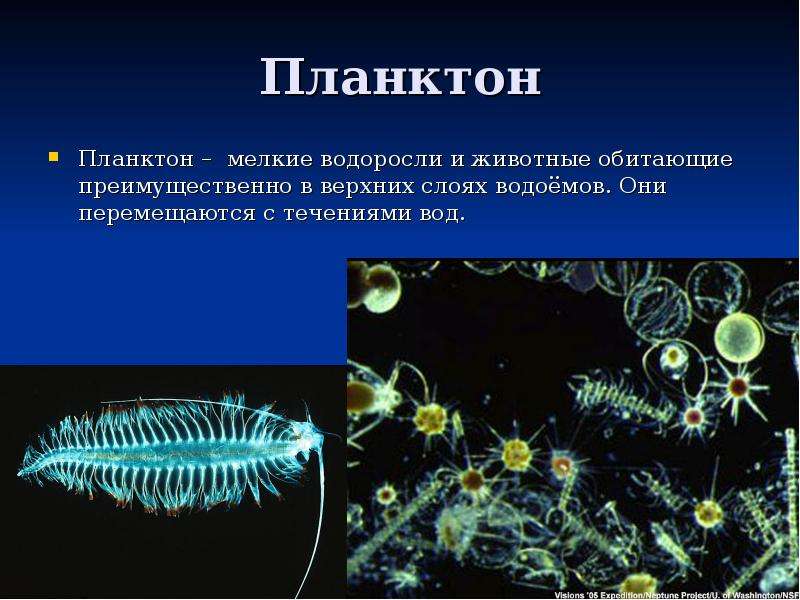 Ж. океаногр. 60, 719–729
Ж. океаногр. 60, 719–729
35 Schippers, P. et al. (2004) Повышение содержания CO в атмосфере
2
способствует
продуктивности фитопланктона. Экол. лат. 7, 446–451
36 Riebesell, U. et al. (2000) Уменьшение кальцификации морского планктона в ответ на
повышенное содержание CO в атмосфере
2
.Nature 407, 634–637
37 Barker, S. et al. (2003) Будущее углеродного цикла: обзор,
реакция кальцификации, балласт и обратная связь по атмосферному CO
2
.
Филос. Транс. Р. Соц. Лонд. сер. A 361, 1977–1999
38 Бойд, П.В. и Дони, С. К. (2003) Влияние изменения климата и
процессов обратной связи на углеродный цикл океана. В Ocean Biogeochem-
istry (Fasham, M.J.R., ed.), стр. 157–193, Springer-Verlag
39 Kurihara, H.и другие. (2004) Сублетальное воздействие повышенной концентрации
CO
2
на планктонных копепод и морских ежей. Ж. океаногр. 60,
743–750
40 Портнер, Х. О. и другие. (2004) Биологическое воздействие повышенных концентраций CO
О. и другие. (2004) Биологическое воздействие повышенных концентраций CO
2
океана: уроки физиологии животных и истории Земли.
Ж. Океан.гр. 60, 705–718
41 Бродгейт, В. (2004) Исследование поверхности океана – нижних слоев атмосферы.
Научный план и стратегия реализации, отчет 50 IGBP (http://
www.uea.ac.uk/env/solas)
42 Hall, J., ed. (2004) IMBER (Интегрированная морская биогеохимия и
исследования экосистем) Научный план и стратегия реализации,
IGBP и SCOR (http://www.imber.info)
43 Horwood, J. et al. (2000) Планктонное определение изменчивости и
устойчивости рыболовства. Дж. Планк. Рез. 22, 1419–1422
44 Ayon, P. et al. (2004) Тенденции объема зоопланктона у побережья Перу между
1964 и 2001 годами. ICES J.мар. 61, 478–484
45 Beaugrand, G. et al. (2003) Влияние планктона на пополнение запасов трески в
Северном море. Nature 426, 661–664
46 Beaugrand, G. and Reid, P.C. (2003) Долгосрочные изменения фито-
and Reid, P.C. (2003) Долгосрочные изменения фито-
планктона, зоопланктона и лосося, связанные с климатом. Глобальное изменение
Биол. 9, 801–817
47 Карл Д.М. и другие. (2001) Долгосрочные изменения в структуре и продуктивности планктонного сообщества
в субтропическом круговороте северной части Тихого океана: гипотеза сдвига домена
.Глубоководные рез. II 48, 1449–1470
48 Бейтс, Н.Р. (2001) Межгодовая изменчивость океанических биогеохимических свойств CO
2
и
в субтропическом
круговороте западной части Северной Атлантики. Глубоководные рез. II 48, 1507–1528
49 Бендер М. и др. (2002) A Крупномасштабный CO
2
План наблюдений: на месте
Океаны и атмосфера (LSCOP), Специальный отчет NOAA OAR
50 Aiken, J. et al. (2000) Атлантический меридиональный разрез: обзор и синтез данных.прог. океаногр. 45, 257–312
51 Гуревич Дж. и Хеджес Л.В. (1999) Статистические вопросы экологического метаанализа
. Ecology 80, 1142–1149
Ecology 80, 1142–1149
52 Micheli, F. (1999) Эвтрофикация, рыболовство и динамика потребительских ресурсов
в морских пелагических экосистемах. Science 285, 1396–1398
53 Ворм, Б. и Майерс, Р.А. (2003) Мета-анализ взаимодействий
трески и креветок показывает нисходящий контроль в океанических пищевых сетях. Экология
84, 162–173
54 Дринкуотер, К.Ф. и др. (2004) Реакция морских экосистем на
изменчивость климата, связанную с Североатлантическим колебанием. В
The North Atlantic Oscillation: Climatic Significance and Environment-
Mental Impact (Hurrell, J.M. et al., eds), pp. 211–234, American
Geophysical Union
55 Holligan, P.M. и другие. (1983) Спутниковые и судовые исследования продукции кокколитофоридов
вдоль края континентального шельфа. Nature 304, 339–342
56 Смит.Т.Дж. и другие. (2004) Временные ряды активности кокколитофорид в
Баренцевом море по спутниковым снимкам за двадцать лет. Геоф. Рез. лат.
Геоф. Рез. лат.
31, doi:10.1029/2004GL019735
57 Beaugrand, G. et al. (2002) Разнообразие каланоидных копепод Севера
Атлантики и прилегающих морей: видовые ассоциации и биогеография.
Мар. Экол. прог. сер. 232, 179–195
58 Fraser, J.H. (1968) История отбора проб планктона. Зоопланктон
Отбор проб (ЮНЕСКО, изд.), стр. 11–18, ЮНЕСКО
59 Wiebe, P.H. и Бенфилд, М.С. (2003) От сети Хенсена к
четырехмерной биологической океанографии. прог. океаногр. 56, 7–136
60 Харди, А.С. (1926) Новый метод исследования планктона. Nature 118,
630
61 Баттен, С.Д. и другие. (2003) Отбор проб CPR: техническая основа,
материалов и методов, согласованность и сопоставимость. прог. океан-
ногр. 58, 193–215
62 Баттен, С.Д.и Уэлч, Д.В. (2003) Изменения
популяций океанического зоопланктона в северо-восточной части Тихого океана, связанные с возможным
изменением климатического режима в 1998/1999 гг. Глубокое море Res. II 51, 863–873
Глубокое море Res. II 51, 863–873
63 Hosie, G.W. и другие. (2002) Развитие Южного океана
Съемка с помощью непрерывного регистратора планктона. прог. океаногр. 58, 263–283
64 Southward, A.J. и другие. (2004) Долгосрочные океанографические и экологические исследования
в западной части Ла-Манша.Доп. Мар биол. 47, 1–104
65 Боград С.Я. и другие. (2003) CalCOFI: полвека физических, химических
и биологических исследований в Калифорнийской текущей системе. Глубоководный
Рез. II 50, 2349–2353
Review TRENDS in Ecology and Evolution Vol.20 No.6 June 2005344
www.sciencedirect.com
Reservoir Nutrient and Plankton Dynamics
Колониальный хризофит рода Synura, этот организм является одним из нескольких таксонов фитопланктона, которые могут вызывать неприятный вкус a Резервуары проводились с октября 1998 г. по сентябрь 1999 г., которые продолжаются, с некоторыми изменениями, по ежеквартальному графику до настоящего времени.
Лабораторные измерения концентрации питательных веществ были выполнены сотрудниками MWRA Центральной лаборатории острова Дир. Пространственный охват программы (шесть станций в Куаббине, четыре станции в Вачусетте, пробы на каждой из которых отбираются на разной глубине) и частота отбора проб в сочетании с эффективностью лабораторных измерений обеспечивают наиболее полную доступную в настоящее время базу данных о динамике биогенных веществ и планктона в обоих водохранилищах. .
Основные находки на водохранилище Куаббин включают:
- Заметные сезонные и вертикальные колебания концентрации питательных веществ, обусловленные динамикой фитопланктона
- Небольшой горизонтальный градиент концентраций кремнезема коррелирует со временем гидравлического пребывания
- Годовой цикл сукцессии и численности фитопланктона, характерный для многих умеренных олиготрофных систем
Основные находки водохранилища Вачусетт включают:
- Заметные сезонные и вертикальные колебания концентрации питательных веществ, обусловленные динамикой фитопланктона
- Межгодовые сдвиги концентраций питательных веществ и интенсивности других параметров, соответствующие разным влияниям переноса Куаббина и водораздела Вачусетт
- Сообщество фитопланктона с преобладанием диатомей и хризофитов, типичное для многих олиготрофных систем с мягкой водой, расположенных в умеренной зоне
- Годовой цикл последовательности и численности фитопланктона, также характерный для этого типа системы, но с дополнительными особенностями, уникальными для водохранилища Вачусетт
В отчете Wachusett также представлен обзор данных о планктоне, полученных независимо из следующих источников:
- Отбор проб DWSP проводится еженедельно (если позволяют ледовые условия) в водозаборе Косгроув с 1987 г.
- Исторические пробы резервуаров, проведенные в 1988, 1989, 1995 и 1996 годах; и
- Исторические данные, собранные MWRA

Кроме того, DCR предоставляет подробную характеристику «перетока» Куаббина, гидродинамического явления, возникающего в результате ежегодного переноса воды из водохранилища Куаббин в Вачусетт через акведук Куаббин.Наконец, этот отчет объединяет исторические записи о питательных веществах и планктоне, а также связанную с ними информацию, полученную в результате многочисленных исследований водохранилища Вачусетт, проведенных консультантами.
Динамика питательных веществ и планктона в водохранилище Куаббин
Динамика питательных веществ и планктона в водохранилище Вачусетт
Промежуточный поток Quabbin — нисходящий поток
Видеорегистратор планктона (VPR) - Океанографический институт Вудс-Хоул
Что это такое и почему мы его используем?
Видеорегистратор планктона (VPR) представляет собой систему подводного видеомикроскопа, которая делает изображения планктона и твердых частиц размером от 50 микрон до нескольких сантиметров.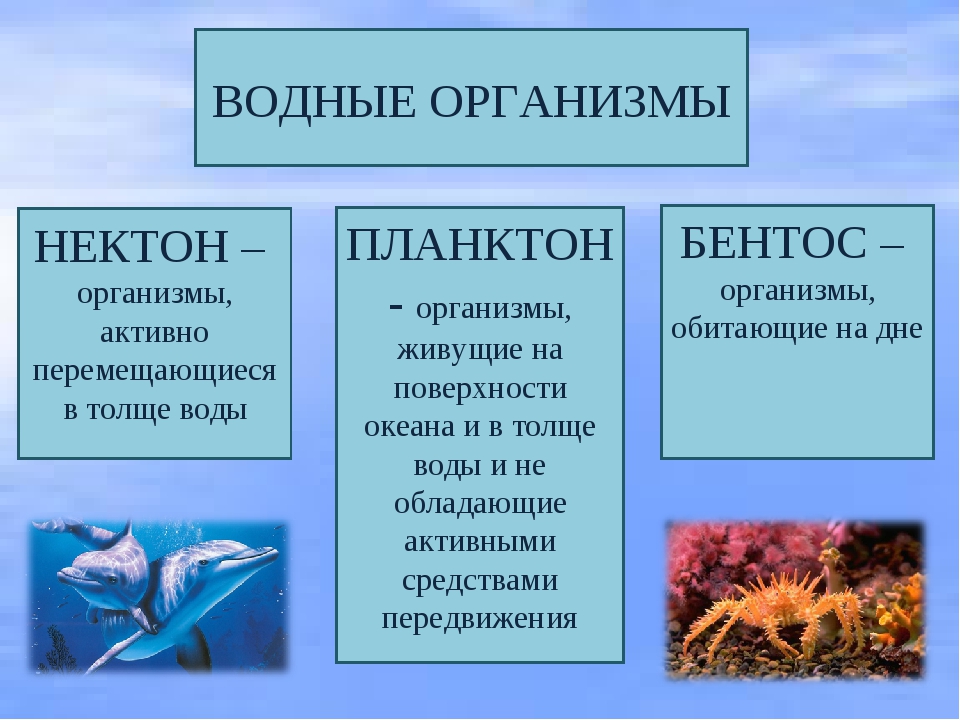 Инструмент используется, чтобы помочь ученым быстро измерить закономерности распределения планктона, не разрушая его тонкие формы, как это может произойти при использовании сетей и бутылок.
Инструмент используется, чтобы помочь ученым быстро измерить закономерности распределения планктона, не разрушая его тонкие формы, как это может произойти при использовании сетей и бутылок.
Большая часть живого вещества в океане — это планктон — мелкие животные, растения и микробы, которые пассивно дрейфуют по течению. Среди них веслоногие, возможно, самые многочисленные животные на Земле и важное звено в пищевой цепи между первичными производителями и более высокими уровнями, такими как рыба. Изучение планктона помогает ученым понять многое другое об океане, например, изменения в рыбных запасах, загрязнение и климат.
Как это работает?
Видеорегистратор планктона (VPR) по сути является подводным микроскопом. Видеокамера, установленная в одном из рукавов, фокусируется на точке посередине между двумя рукавами. Стробоскоп на другом плече освещает отображаемый объем и мигает 60 раз в секунду, создавая 60 изображений частиц и планктона в воде в секунду. Затем изображения сохраняются на жестком диске компьютера, а затем наносятся на график.
Прибор монтируется в раму и спускается на воду с кормы корабля.Иногда CTD также устанавливается рядом с VPR для сбора информации о глубине, температуре и солености одновременно с каждым видеоизображением. Прибор опускают под воду на максимальную глубину 350 метров, чтобы мы получили профиль содержания и типа планктона/частиц, а также температуры и солености.
Недавно был разработан быстрый 3-осевой VPR для скорости буксировки до 12 узлов. Он может обрабатывать автономные волнообразные и боковые движения. Этот быстрый VPR использовался в августе 2003 г. на НИС Knorr для проведения непрерывной трансатлантической съемки протяженностью 6000 км со скоростью 12 узлов с волнами через каждые 1 км до глубины 120 м.
Какие платформы нужны?
Использовался на:
- Буксируемые корпуса (депрессор с V-образным плавником, Seasoar и новый Fast towfish)
- ROV (JASON и SeaRover)
- АНПА (РЕМУС)
- автономный профилирующий швартовщик
Ограничения:
- в настоящее время не может идентифицировать планктон до видового уровня. 90 324 недовыборки редких таксонов (например, <50/м3).

Видео, демонстрирующее операции VPR в западной части Северной Атлантики.
HWS: Институт Фингер-Лейкс
Планы уроков SOS/рабочие листы для учащихся
Руководство для учителей по планктону — основным производителям водного мира
Введение в урок
Планктон находится в нижней части пищевой цепи в водных системах. Без них не было бы жизни ни в озерах, ни в прудах, ни в океанах. Проще говоря, фитопланктон является первичным производителем или микроскопической растительной жизнью водной системы, в то время как зоопланктон, следующий по цепочке, является их потребителем.Слово «планктон» происходит от греческого слова «планктос», означающего «бродяга». Большинство планктона не умеют плавать, они свободно плавают в водных системах во власти приливов, волн и течений. Однако многие планктоны обладают необычной симметрией или приспособлениями, которые позволяют им избегать нападения хищников, плавать, не погружаясь глубже в толщу воды, где меньше солнечного света, и обеспечивать некоторую подвижность. Динофлагелляты, например, имеют два жгутика или хвоста, что ограничивает их подвижность в воде.
Проще говоря, фитопланктон является первичным производителем или микроскопической растительной жизнью водной системы, в то время как зоопланктон, следующий по цепочке, является их потребителем.Слово «планктон» происходит от греческого слова «планктос», означающего «бродяга». Большинство планктона не умеют плавать, они свободно плавают в водных системах во власти приливов, волн и течений. Однако многие планктоны обладают необычной симметрией или приспособлениями, которые позволяют им избегать нападения хищников, плавать, не погружаясь глубже в толщу воды, где меньше солнечного света, и обеспечивать некоторую подвижность. Динофлагелляты, например, имеют два жгутика или хвоста, что ограничивает их подвижность в воде.
Будучи основными производителями в водном мире, фитопланктон использует для своей энергии свет. Они собирают энергию солнечного света, преобразуют эту энергию в углеводы и выделяют кислород. Им также нужны питательные вещества. Подобно наземным растениям, которые получают питательные вещества из почвы, фитопланктон поглощает питательные вещества из толщи воды через свои клеточные стенки. Азот и фосфор являются основными питательными веществами, потребляемыми фитопланктоном, хотя эти организмы используют гораздо больше. Фосфор является основным питательным веществом, ограничивающим рост растений в водных системах, в то время как азот является основным питательным веществом, ограничивающим рост наземных растений.
Азот и фосфор являются основными питательными веществами, потребляемыми фитопланктоном, хотя эти организмы используют гораздо больше. Фосфор является основным питательным веществом, ограничивающим рост растений в водных системах, в то время как азот является основным питательным веществом, ограничивающим рост наземных растений.
Как и наземные растения, фитопланктон играет важную роль в производстве кислорода. Днем во время фотосинтеза фитопланктон выделяет кислород, а ночью во время дыхания расходует кислород.
Формулы фотосинтеза и дыхания:
Фотосинтез: 6CO2 + Esun + 6h3O 6O2 + C6 h22O6
Дыхание: 6O2 + C6 h22 O6 6 CO2 + Ecells + 6 h3O
В здоровых водных системах количество кислорода, вырабатываемого фитопланктоном в течение дня, компенсирует потери кислорода ночью, когда они не фотосинтезируют.Этот цикл может стать разрушительным, если процессы дыхания потребляют слишком много кислорода, вызывая стресс у других водных организмов. Кислородное истощение может иметь место в озерах или прудах, которые считаются эвтрофными или чрезмерно продуктивными и богатыми питательными веществами. Ученые определяют скорость фотосинтеза и дыхания в водных системах, чтобы увидеть, находятся ли они в равновесии. Один из способов сделать это — измерить скорость производства и потребления кислорода в образце воды из озера или пруда, который вы изучаете.С помощью расчетов вы можете определить чистую производительность кислорода в системе.
Ученые определяют скорость фотосинтеза и дыхания в водных системах, чтобы увидеть, находятся ли они в равновесии. Один из способов сделать это — измерить скорость производства и потребления кислорода в образце воды из озера или пруда, который вы изучаете.С помощью расчетов вы можете определить чистую производительность кислорода в системе.
Цели урока
На этом уроке учащиеся узнают, как готовить и брать пробы воды с разных глубин озера или пруда и измерять количество растворенного кислорода в каждой пробе. Затем они рассчитают чистую продуктивность кислорода и ответят на вопросы, представленные в Упражнении 1. Во время Упражнения 2 учащимся будет предложено выдвинуть гипотезу о влиянии удобрений на фитопланктон из проб воды путем измерения мутности воды. образцы.
Стандарты MST:
- Стандарт 1: Основная идея 1
- Стандарт 4: Основная идея 6
Виды деятельности
Мероприятие 1. Измерение первичной производительности
Мероприятие 2 - Как удобрения, такие как фосфорные, влияют на рост фитопланктона?
границ | Классификация глубокого обучения озерного зоопланктона
1.
 Введение
Введение Планктон — ключевой компонент биосферы Земли. К ним относятся все водные организмы, дрейфующие по течению, от мельчайших бактерий и микроводорослей до личинок позвоночных и беспозвоночных.Фотосинтетический фитопланктон отвечает примерно за половину глобальной первичной продукции (Behrenfeld et al., 2001) и, следовательно, играет центральную роль в фиксации углерода в атмосфере и производстве кислорода. Зоопланктон представляет собой обширную группу водных микроорганизмов, насчитывающую более десятков тысяч видов (Sournia et al., 1991) и включающую как плотоядных, так и травоядных, причем последние питаются фитопланктоном. Планктон является важнейшим компонентом водных пищевых цепей, производя органическое вещество, которое является основным источником массы и энергии для более высоких трофических уровней (Lotze et al., 2019), и служат пищей личинкам рыб (Banse, 1995). Смерть и экскреция планктонных организмов приводит к секвестрации огромного количества углерода, что регулирует биологический углеродный насос локально и глобально (Volk and Hoffert, 1985). Таким образом, биоразнообразие и динамика планктона напрямую влияют на климат, рыболовство и средства к существованию человеческого населения вблизи водоемов.
Таким образом, биоразнообразие и динамика планктона напрямую влияют на климат, рыболовство и средства к существованию человеческого населения вблизи водоемов.
Планктонные организмы, в основном небольшие по размеру, имеют короткую продолжительность жизни и высокую чувствительность к условиям окружающей среды, что делает их разнообразие и численность очень эффективными индикаторами изменений окружающей среды и здоровья экосистемы.Особенно в пресноводных экосистемах они страдают от комбинированного воздействия местного воздействия человека и глобальных изменений, таких как потепление и инвазивные виды (Williamson et al., 2009). Информация об отдельных видах планктона также имеет решающее значение для мониторинга вредоносного цветения водорослей, которое может нанести огромный экологический и экономический ущерб и иметь серьезные последствия для здоровья населения (Huisman et al., 2018). Разнообразие и численность планктона обычно измеряются с помощью трудоемких методов отбора проб и микроскопии, которые имеют ряд ограничений, таких как высокая стоимость, специализированный персонал, низкая производительность, длительное время обработки проб, субъективность классификации и низкая прослеживаемость и воспроизводимость данных. .Эти ограничения стимулировали разработку множества альтернативных и автоматизированных инструментов мониторинга планктона (Lombard et al., 2019), некоторые из которых недавно применялись в пресноводных системах (Spanbauer et al., 2020; Merz et al., 2021; Tapics и др., 2021). Недавно разработанные методы, такие как эДНК, открывают большие перспективы, в частности, для мониторинга биоразнообразия в больших пространственных и временных масштабах, для идентификации загадочных видов (не поддающихся обнаружению морфологически) и для учета генетического/функционального разнообразия (Deiner et al., 2017), но еще не реализованы для высокочастотного мониторинга на месте.
.Эти ограничения стимулировали разработку множества альтернативных и автоматизированных инструментов мониторинга планктона (Lombard et al., 2019), некоторые из которых недавно применялись в пресноводных системах (Spanbauer et al., 2020; Merz et al., 2021; Tapics и др., 2021). Недавно разработанные методы, такие как эДНК, открывают большие перспективы, в частности, для мониторинга биоразнообразия в больших пространственных и временных масштабах, для идентификации загадочных видов (не поддающихся обнаружению морфологически) и для учета генетического/функционального разнообразия (Deiner et al., 2017), но еще не реализованы для высокочастотного мониторинга на месте.
Если, с одной стороны, изучение пресноводной среды дает возможность подойти к нескольким вопросам, связанным с (i) автоматическим распознаванием таксонов планктона в системах, качество воды которых строго контролируется, и (ii) созданием временных рядов популяций планктона, полезных для как исследования, так и управление озером, с другой стороны, это дает ряд практических преимуществ. Количество видов, присутствующих в озере, составляет порядка нескольких сотен, а состав сообщества меняется в масштабе десятилетий (Pomati et al., 2012), и практически все озера одного и того же региона, как правило, относятся к одному и тому же географическому/климатическому региону и к одному и тому же видовому пулу таксонов планктона (Monchamp et al., 2019). Это позволило бы нам обрабатывать данные о реальных озерах с меньшей необходимостью учитывать изменчивость видов, довольно быстро создать базу данных, включающую все наблюдаемые таксоны, и легко использовать наши модели для более чем одного участка. Кроме того, для озер обычно характерны более низкие уровни непланктонных взвешенных веществ (например, песка, мусора) по сравнению с прибрежной морской средой, поэтому можно ожидать работы с более чистыми изображениями, с относительно небольшим количеством небиологических или небиологических обнаруживаются узнаваемые объекты.
Количество видов, присутствующих в озере, составляет порядка нескольких сотен, а состав сообщества меняется в масштабе десятилетий (Pomati et al., 2012), и практически все озера одного и того же региона, как правило, относятся к одному и тому же географическому/климатическому региону и к одному и тому же видовому пулу таксонов планктона (Monchamp et al., 2019). Это позволило бы нам обрабатывать данные о реальных озерах с меньшей необходимостью учитывать изменчивость видов, довольно быстро создать базу данных, включающую все наблюдаемые таксоны, и легко использовать наши модели для более чем одного участка. Кроме того, для озер обычно характерны более низкие уровни непланктонных взвешенных веществ (например, песка, мусора) по сравнению с прибрежной морской средой, поэтому можно ожидать работы с более чистыми изображениями, с относительно небольшим количеством небиологических или небиологических обнаруживаются узнаваемые объекты.
Среди автоматизированных подходов к мониторингу планктона методы визуализации обладают наибольшим потенциалом для получения стандартизированной и воспроизводимой количественной оценки численности, биомассы, разнообразия и морфологии планктона в разных масштабах (Lombard et al. , 2019; Merz et al., 2021). В настоящее время существует несколько устройств цифровой обработки изображений in-situ , таких как Imaging FlowCytobot (Olson and Sosik, 2007), камера Scripps Plankton Camera (SPC) (Orenstein et al., 2020), Video Plankton Recorder (Davis, 1992), SIPPER. и модифицированный SPC с двойным увеличением (www.aquascope.ch) (Merz et al., 2021).
, 2019; Merz et al., 2021). В настоящее время существует несколько устройств цифровой обработки изображений in-situ , таких как Imaging FlowCytobot (Olson and Sosik, 2007), камера Scripps Plankton Camera (SPC) (Orenstein et al., 2020), Video Plankton Recorder (Davis, 1992), SIPPER. и модифицированный SPC с двойным увеличением (www.aquascope.ch) (Merz et al., 2021).
Эти системы цифровой обработки изображений могут создавать очень большие объемы изображений планктона, особенно если они развернуты на месте для непрерывного автоматизированного мониторинга (Orenstein et al., 2020; Merz et al., 2021). Хотя выделение признаков изображения, описывающих важные характеристики планктона, такие как размер и форма, хорошо зарекомендовало себя (Orenstein et al., 2020; Merz et al., 2021), классификация больших объемов объектов по различным таксономическим категориям планктона по-прежнему остается сложной задачей. и представляет собой наиболее важный компонент для мониторинга планктона (MacLeod et al. , 2010). Автоматизированная классификация объектов планктона на изображениях может помочь систематикам аннотировать изображения и позволить проводить выборку и подсчет таксонов с высоким временным и пространственным разрешением. Автоматизация мониторинга планктона может стать ключевой инновацией в области оценки и управления качеством воды, водным биоразнообразием, инвазивными видами, влияющими на экосистемные услуги (например, паразиты, инвазивные мидии), и ранним предупреждением о вредоносном цветении водорослей.
, 2010). Автоматизированная классификация объектов планктона на изображениях может помочь систематикам аннотировать изображения и позволить проводить выборку и подсчет таксонов с высоким временным и пространственным разрешением. Автоматизация мониторинга планктона может стать ключевой инновацией в области оценки и управления качеством воды, водным биоразнообразием, инвазивными видами, влияющими на экосистемные услуги (например, паразиты, инвазивные мидии), и ранним предупреждением о вредоносном цветении водорослей.
Автоматизированная классификация планктона характеризуется рядом особенностей, которые делают эту задачу менее простой, чем другие подобные задачи.Наборы данных, используемые для обучения, а также изображения, проанализированные после развертывания, охватывают широкие таксономические диапазоны, которые очень неравномерно распределены (некоторые таксоны встречаются очень часто, а другие встречаются редко — это называется дисбалансом данных или дисбалансом классов ). (Orenstein et al., 2015), и это распределение меняется со временем, например, с появлением или исчезновением новых таксонов или с изменением стадии жизни вида, доминирующего в сигнале (Schröder et al., 2018). Более того, многие изображения не принадлежат ни к одному таксону (например,г., грязь), либо их невозможно идентифицировать из-за низкого разрешения, положения, фокусировки или кадрирования. Кроме того, маркировка этих наборов данных требует больших усилий, поскольку они должны быть аннотированы экспертами-таксономистами, а выборка изображений из видео, как это делается, например, для фотоловушек (Tabak et al., 2019), бесполезна, поскольку выравнивание организмов по отношению к камере обычно не меняется в течение всего времени экспозиции.
(Orenstein et al., 2015), и это распределение меняется со временем, например, с появлением или исчезновением новых таксонов или с изменением стадии жизни вида, доминирующего в сигнале (Schröder et al., 2018). Более того, многие изображения не принадлежат ни к одному таксону (например,г., грязь), либо их невозможно идентифицировать из-за низкого разрешения, положения, фокусировки или кадрирования. Кроме того, маркировка этих наборов данных требует больших усилий, поскольку они должны быть аннотированы экспертами-таксономистами, а выборка изображений из видео, как это делается, например, для фотоловушек (Tabak et al., 2019), бесполезна, поскольку выравнивание организмов по отношению к камере обычно не меняется в течение всего времени экспозиции.
Модели классификации изображений делятся на несколько широких категорий, включая неконтролируемые модели (которые группируют и классифицируют изображения без каких-либо тегов, назначаемых вручную), контролируемые модели (которые используют обучающую библиотеку изображений, идентифицированных вручную, для разработки модели классификации) и гибридные модели ( которые сочетают в себе аспекты контролируемого и неконтролируемого обучения). Несмотря на то, что в настоящее время проводятся исследования, основанные на обучении без учителя (Salvesen et al., 2020; Schröder et al., 2020) или на разработке определенных видов предварительной обработки данных (Zhao et al., 2010; Zheng et al., 2017). ), текущий уровень техники для классификации наборов данных о планктоне чаще всего включает глубокие сверточные нейронные сети, обученные на классифицированных вручную изображениях (Dai et al., 2016; Dai et al., 2017; Lee et al., 2016; Li and Cui). , 2016; Py et al., 2016; Orenstein and Beijbom, 2017; Cui et al., 2018; Дункер и др., 2018; Луо и др., 2018; Родригес. и др., 2018; Бочински и др., 2019; Люмини и Нанни, 2019 г.; Эрола и др., 2020 г.; Керр и др., 2020 г.; Люмини и др., 2020 г.; Го и др., 2021 г.; Henrichs et al., 2021), которые обеспечивают большую гибкость между приложениями и продемонстрировали более удовлетворительные результаты, чем ручное извлечение признаков (González et al., 2019). Эти приложения очень часто прибегают к трансферному обучению (Tan et al.
Несмотря на то, что в настоящее время проводятся исследования, основанные на обучении без учителя (Salvesen et al., 2020; Schröder et al., 2020) или на разработке определенных видов предварительной обработки данных (Zhao et al., 2010; Zheng et al., 2017). ), текущий уровень техники для классификации наборов данных о планктоне чаще всего включает глубокие сверточные нейронные сети, обученные на классифицированных вручную изображениях (Dai et al., 2016; Dai et al., 2017; Lee et al., 2016; Li and Cui). , 2016; Py et al., 2016; Orenstein and Beijbom, 2017; Cui et al., 2018; Дункер и др., 2018; Луо и др., 2018; Родригес. и др., 2018; Бочински и др., 2019; Люмини и Нанни, 2019 г.; Эрола и др., 2020 г.; Керр и др., 2020 г.; Люмини и др., 2020 г.; Го и др., 2021 г.; Henrichs et al., 2021), которые обеспечивают большую гибкость между приложениями и продемонстрировали более удовлетворительные результаты, чем ручное извлечение признаков (González et al., 2019). Эти приложения очень часто прибегают к трансферному обучению (Tan et al. , 2018), которое заключается в использовании моделей, предварительно обученных на большом наборе данных изображений [обычно ImageNet (Deng et al., 2009)] и их адаптация к конкретной задаче распознавания изображений. Трансферное обучение требует сравнительно меньше аннотированных человеком данных в целевой области, чтобы получить разумную модель после обучения, чем модель, обученная с нуля. Это также ускоряет процесс обучения и приводит к повышению производительности модели. Трансферное обучение использовалось в двухэтапном процессе для устранения дисбаланса данных (Lee et al., 2016), но чаще всего оно используется, поскольку позволяет обучать очень большие модели в разумные сроки.Основные различия в различных приложениях к планктону часто заключаются в способе предварительной обработки изображений. Например, Дай и др. (2017) фильтрует изображения по-разному и подает как исходные, так и отфильтрованные изображения в качестве входных данных для моделей, Cui et al. (2018) применяет логарифмическое улучшение изображения к черно-белым изображениям, а Люмини и Нанни (2019) тестируют различные способы изменения размера изображений.
, 2018), которое заключается в использовании моделей, предварительно обученных на большом наборе данных изображений [обычно ImageNet (Deng et al., 2009)] и их адаптация к конкретной задаче распознавания изображений. Трансферное обучение требует сравнительно меньше аннотированных человеком данных в целевой области, чтобы получить разумную модель после обучения, чем модель, обученная с нуля. Это также ускоряет процесс обучения и приводит к повышению производительности модели. Трансферное обучение использовалось в двухэтапном процессе для устранения дисбаланса данных (Lee et al., 2016), но чаще всего оно используется, поскольку позволяет обучать очень большие модели в разумные сроки.Основные различия в различных приложениях к планктону часто заключаются в способе предварительной обработки изображений. Например, Дай и др. (2017) фильтрует изображения по-разному и подает как исходные, так и отфильтрованные изображения в качестве входных данных для моделей, Cui et al. (2018) применяет логарифмическое улучшение изображения к черно-белым изображениям, а Люмини и Нанни (2019) тестируют различные способы изменения размера изображений.
Кроме того, несколько моделей можно использовать совместно для повышения производительности (будь то устранение дисбаланса данных или достижение более высокой взвешенной точности).Двумя основными подходами к объединению нескольких моделей являются совместные модели и ансамбли. Первый состоит из обучения моделей вместе для получения общего результата (Dai et al., 2017; Kerr et al., 2020), а второй обучает модели отдельно и объединяет результаты на более позднем этапе. Недавно для противодействия дисбалансу данных использовались совместные модели, обеспечивающие высокие характеристики одноканальных (то есть черно-белых) изображений, полученных на станции L4 в западной части Ла-Манша (Kerr et al., 2020). Однако это предполагает одновременное развертывание нескольких моделей, что приводит к очень высокому использованию памяти, если только не используются меньшие версии обычно используемых моделей (таким образом, не допуская переноса обучения). Объединение позволяет объединить практически любое количество учащихся и привело к очень удовлетворительным результатам при объединении различных архитектур (где DenseNets чаще всего работают лучше всего) или видов предварительной обработки (Lumini and Nanni, 2019).
Упомянутые методы автоматической классификации планктона применялись главным образом в прибрежных местообитаниях с соленой водой.Насколько нам известно, единственная предыдущая работа по классификации изображений пресной воды была проведена Hong et al. (2020), где данные не поступают из автоматизированной системы, и они изучают небольшой сбалансированный набор данных, отсортированный по четырем категориям (дафния, каланоид, самка циклопоида, самец циклопоида), и получают максимальную точность классификации 93%.
В этой статье мы изучаем классификацию планктонных организмов из озерных экосистем на новом наборе данных изображений озерного планктона, которые мы предоставляем в свободный доступ, вместе с кодом, который позволяет легко обучать и развертывать наши глубокие нейронные сети.Мы анализируем изображения планктона, полученные с помощью камеры Scripps Plankton Camera (DSPC) с двойным увеличением, которая представляет собой микроскоп для получения изображений в темном поле, который в настоящее время используется в озере Грайфензее (Швейцария) (Merz et al. , 2021), и, в частности, изображения с 0,5-кратным увеличением. , который нацелен на зоопланктон и крупные колониеобразующие таксоны фитопланктона в диапазоне от 100 мкм до 1 см. Мы вручную аннотировали 17943 изображения, состоящие из n c = 35 неравномерно распределенных категорий (классов), которые были собраны in-situ с использованием DSPC, установленного на глубине 3 м в озере Грайфензее.Мы предлагаем набор моделей глубокого обучения, использующих трансферное обучение, и объединяем их с помощью версий совместного и ансамблевого обучения. В частности, мы изучаем несколько способов объединения наших моделей на основе недавних статистических данных (D’Ascoli et al., 2020; Geiger et al., 2020). Мы оцениваем производительность наших моделей на общедоступных наборах данных, получая небольшое, но систематическое увеличение производительности по сравнению с предыдущей литературой. Простейшие из представленных моделей использовались для анализа части данных в работе Merz et al.
, 2021), и, в частности, изображения с 0,5-кратным увеличением. , который нацелен на зоопланктон и крупные колониеобразующие таксоны фитопланктона в диапазоне от 100 мкм до 1 см. Мы вручную аннотировали 17943 изображения, состоящие из n c = 35 неравномерно распределенных категорий (классов), которые были собраны in-situ с использованием DSPC, установленного на глубине 3 м в озере Грайфензее.Мы предлагаем набор моделей глубокого обучения, использующих трансферное обучение, и объединяем их с помощью версий совместного и ансамблевого обучения. В частности, мы изучаем несколько способов объединения наших моделей на основе недавних статистических данных (D’Ascoli et al., 2020; Geiger et al., 2020). Мы оцениваем производительность наших моделей на общедоступных наборах данных, получая небольшое, но систематическое увеличение производительности по сравнению с предыдущей литературой. Простейшие из представленных моделей использовались для анализа части данных в работе Merz et al. (2021).
(2021).
2. Материалы и методы
2.1. Сбор данных
Мы использовали изображения, полученные с помощью DSPC (Merz et al., 2021), развернутого в озере Грайфензее, и полученные от таксонов дикого планктона в период с 2018 по 2020 год. DSPC делает изображения микроскопических таксонов планктона с заданной пользователем частотой и частотой. интервалы времени (подробнее и настройки камеры см. в Merz et al., 2021). Исходные полнокадровые изображения могут содержать от нуля до нескольких изображений планктонных организмов, а также неорганического вещества.Полные кадры сегментируются на месте в режиме реального времени, а области интереса (ROI), которые содержат, например, планктонные организмы, сохраняются и используются для выделения и классификации признаков изображения. Изображения объектов на границе поля зрения камеры получаются кадрированными, но мы их все равно сохраняем, так как большую часть времени мы все еще в состоянии их идентифицировать. Изображения имеют черный фон, что способствует обнаружению ROI. Они имеют разные размеры в зависимости от размера обнаруженного объекта.Для каждой ROI мы выделили 64 морфологических и цветовых признака и выполнили серию графических операций, чтобы сделать изображение более четким. На рисунке 1А мы показываем несколько примеров того, как выглядят окончательные изображения. В дополнительном разделе 1 мы приводим подробное описание набора данных и всех его классов вместе с одним образцом изображения из каждого класса на дополнительном рисунке 1. В дополнительном разделе 2 мы описываем вышеупомянутые 64 морфологических признака.
Они имеют разные размеры в зависимости от размера обнаруженного объекта.Для каждой ROI мы выделили 64 морфологических и цветовых признака и выполнили серию графических операций, чтобы сделать изображение более четким. На рисунке 1А мы показываем несколько примеров того, как выглядят окончательные изображения. В дополнительном разделе 1 мы приводим подробное описание набора данных и всех его классов вместе с одним образцом изображения из каждого класса на дополнительном рисунке 1. В дополнительном разделе 2 мы описываем вышеупомянутые 64 морфологических признака.
Рис. 1.(A) Образцы изображений из DSPC в озере Грайфензее. (B) Изобилие каждого класса в нашем наборе данных. Слово класс предназначено в классификационном смысле и не указывает на таксономический ранг. Обратите внимание, что ось y имеет логарифмическую шкалу.
2.2. Подготовка данных
DSPC можно запускать с двумя разными увеличениями (Merz et al. , 2021), но в этой статье мы сообщаем только об изображениях, сделанных при меньшем увеличении, которые содержат в основном таксоны зоопланктона и несколько крупных колониальных фитопланктона.Мы вручную аннотировали набор данных из 17 943 изображений отдельных объектов в 90 463 n 90 464 90 485 c 90 488 = 35 классов. На рисунке 1B показаны названия всех классов n c вместе с количеством помеченных изображений каждого класса. Обратите внимание, что аннотированных изображений самого распространенного класса (динобрионов) в 300 раз больше, чем самого редкого класса (чаоборус).
, 2021), но в этой статье мы сообщаем только об изображениях, сделанных при меньшем увеличении, которые содержат в основном таксоны зоопланктона и несколько крупных колониальных фитопланктона.Мы вручную аннотировали набор данных из 17 943 изображений отдельных объектов в 90 463 n 90 464 90 485 c 90 488 = 35 классов. На рисунке 1B показаны названия всех классов n c вместе с количеством помеченных изображений каждого класса. Обратите внимание, что аннотированных изображений самого распространенного класса (динобрионов) в 300 раз больше, чем самого редкого класса (чаоборус).
2.3. Открытый доступ к нашему набору данных
Мы называем ZooLake описанный набор данных помеченных изображений планктона.Мы даем подробные сведения о ZooLake в дополнительных разделах 1, 2 и сделали данные общедоступными в Интернете по следующей ссылке: https://data.eawag.ch/dataset/deep-learning-classification-of-zooplankton-from- озера.
2.4. Дальнейшая подготовка данных
Поскольку для большинства моделей глубокого обучения неудобно иметь изображения разного размера, мы изменили размеры наших изображений таким образом, чтобы все они имели одинаковый размер. Два самых простых способа сделать это: (i) изменить размер всех изображений до 128 × 128 пикселей независимо от их начальных размеров, не сохраняя исходные пропорции, или (ii) уменьшить их таким образом, чтобы наибольший размер был равен большинство 128 пикселей (уменьшение изображения не выполняется, если изображение уже меньше) и заполнение их черным фоном, чтобы сделать их 128 × 128.Недостатком первого метода является несоблюдение пропорций. У последнего есть проблема, заключающаяся в том, что в изображениях с очень большим соотношением сторон происходит потеря информации по наименьшему измерению. Два метода сравниваются в Lumini and Nanni (2019), где видно, что процедура (i) дает несколько лучшие результаты в большинстве наборов данных. Кроме того, информацию, потерянную при изменении пропорций объектов, можно восстановить, используя начальное соотношение сторон (и аналогичные величины) в качестве дополнительной входной характеристики.По этим причинам все результаты, которые мы показываем в основном тексте, получены с помощью метода (i).
Два самых простых способа сделать это: (i) изменить размер всех изображений до 128 × 128 пикселей независимо от их начальных размеров, не сохраняя исходные пропорции, или (ii) уменьшить их таким образом, чтобы наибольший размер был равен большинство 128 пикселей (уменьшение изображения не выполняется, если изображение уже меньше) и заполнение их черным фоном, чтобы сделать их 128 × 128.Недостатком первого метода является несоблюдение пропорций. У последнего есть проблема, заключающаяся в том, что в изображениях с очень большим соотношением сторон происходит потеря информации по наименьшему измерению. Два метода сравниваются в Lumini and Nanni (2019), где видно, что процедура (i) дает несколько лучшие результаты в большинстве наборов данных. Кроме того, информацию, потерянную при изменении пропорций объектов, можно восстановить, используя начальное соотношение сторон (и аналогичные величины) в качестве дополнительной входной характеристики.По этим причинам все результаты, которые мы показываем в основном тексте, получены с помощью метода (i).
Чтобы искусственно увеличить количество обучающих изображений, мы использовали метод увеличения данных путем применения случайных деформаций к обучающим изображениям (Abadi et al., 2016). Примененные нами преобразования, которые не изменили распределение данных, включают повороты на 180°, перевороты, масштабирование до 20% и сдвиг до 10%. Что касается морфологических и цветовых признаков, мы рассчитали 44 дополнительных на необработанных изображениях (см. Дополнительный раздел 3) и стандартизировали полученные 111 признаков, чтобы они имели нулевое среднее значение и единичное стандартное отклонение.
2.5. Обучение, проверка и тестирование
Мы разделили наши изображения на наборы для обучения, проверки и тестирования в соотношении 70:15:15. Все сплиты имели распределение классов, аналогичное общему распределению данных. Для всех моделей использовались одни и те же расщепления. Проверочный набор использовался для выбора наилучших (гипер)параметров модели, в то время как тестовый набор откладывался на протяжении всего процесса и использовался только в самом конце для оценки и сравнения производительности всех предложенных моделей.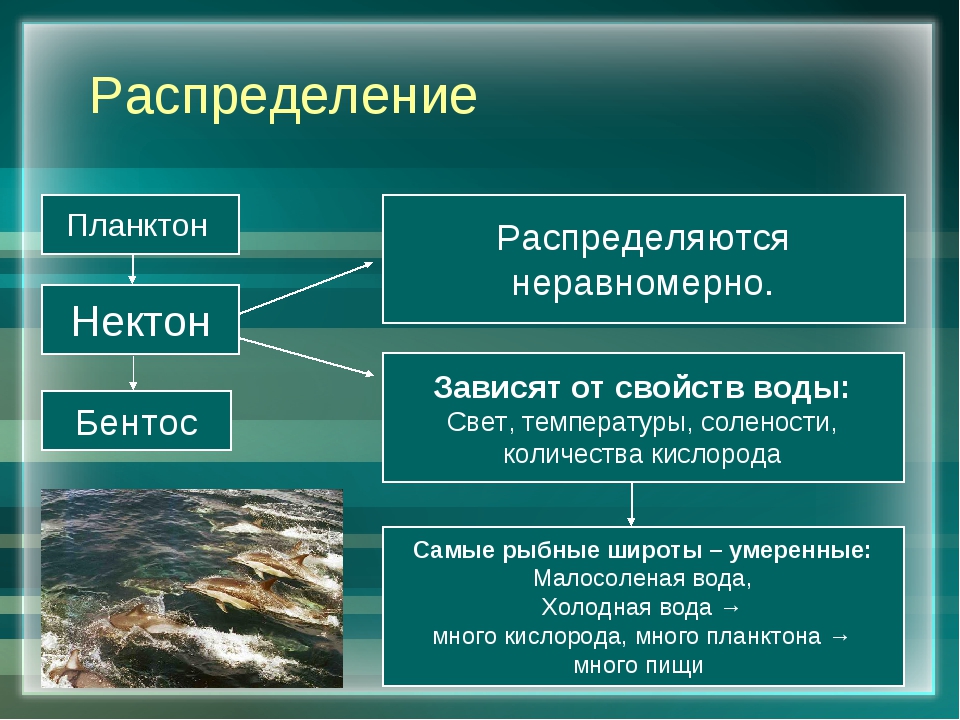
2.5.1. Показатели производительности
Чтобы оценить производительность наших моделей, мы использовали точность, точность, полноту и F1-оценку. В зависимости от конкретного приложения может быть интересна та или иная метрика. В этом разделе мы определяем и кратко объясняем каждый из них с точки зрения истинно положительных результатов (TP), ложноположительных результатов (FP) и ложноотрицательных результатов (FN).
Точность. Точность, A , указывает количество правильных догадок из общего числа изображений,
A=общее количество TPtotal # изображений.(1)Мы рассчитали точность по всему набору данных без выделения классов. Это означает, что в точности преобладают самые современные классы.
Точность. Когда у нас есть набор изображений, которые были отнесены нашими моделями к классу и , нам может быть интересно узнать, сколько из них, как мы ожидаем, на самом деле принадлежат классу и . Для этого мы используем точность P , которая определяется как
.
Сначала мы измеряем точность, относящуюся к каждому отдельному классу, а затем усредняем точность для каждого класса.Это называется макрос средним, и он дает каждой категории одинаковый вес. Это гарантирует, что в этой метрике не преобладают самые распространенные классы.
Отзыв. Отзыв, R , относящийся к классу i , представляет собой долю изображений, принадлежащих к классу i , которые были правильно помечены,
Также в случае отзыва мы используем средние макроэкономические значения.
F1-Оценка. F1-Score объединяет сообщения о точности и отзыве в одно число, которое представляет собой гармоническое среднее между ними:
. Оценка F1=2PRR+P (4)Чтобы получить высокий балл F1 для определенного класса, прогнозы классификатора должны иметь как высокую точность, так и полноту (т.д., низкое количество FP и FN). Также для оценки F1 мы сообщаем средние макроэкономические показатели.
2.6. Архитектуры глубокого обучения
Общая проблема при выборе архитектуры глубокого обучения заключается в том, как наилучшим образом масштабировать архитектуру по глубине, ширине и разрешению изображения. Недавнее решение было дано Таном и Ле (2019), в котором предлагается форма масштабирования для этих трех переменных одновременно вместе с базовой моделью под названием EfficientNetB0, для которой это масштабирование особенно эффективно.Это приводит к более высоким характеристикам, чем предыдущие современные модели, с меньшими затратами с точки зрения параметров модели и количества операций. Предоставленная форма масштабирования позволяет нам получать эффективно масштабируемые модели в зависимости от того, сколько вычислительных ресурсов мы готовы инвестировать. Эти модели, упорядоченные по возрастанию размера, называются EfficientNetB1, EfficientNetB2, EfficientNetB3, EfficientNetB4, EfficientNetB5, EfficientNetB6 и EfficientNetB7. Учитывая вышеупомянутые большие усилия по применению моделей глубокого обучения для классификации планктона, мы считаем, что стоит оценить эффективность этих архитектур в распознавании планктона.Помимо них, мы также тестируем другие архитектуры глубоких нейронных сетей, некоторые из которых уже успешно использовались для решения наших задач.
Недавнее решение было дано Таном и Ле (2019), в котором предлагается форма масштабирования для этих трех переменных одновременно вместе с базовой моделью под названием EfficientNetB0, для которой это масштабирование особенно эффективно.Это приводит к более высоким характеристикам, чем предыдущие современные модели, с меньшими затратами с точки зрения параметров модели и количества операций. Предоставленная форма масштабирования позволяет нам получать эффективно масштабируемые модели в зависимости от того, сколько вычислительных ресурсов мы готовы инвестировать. Эти модели, упорядоченные по возрастанию размера, называются EfficientNetB1, EfficientNetB2, EfficientNetB3, EfficientNetB4, EfficientNetB5, EfficientNetB6 и EfficientNetB7. Учитывая вышеупомянутые большие усилия по применению моделей глубокого обучения для классификации планктона, мы считаем, что стоит оценить эффективность этих архитектур в распознавании планктона.Помимо них, мы также тестируем другие архитектуры глубоких нейронных сетей, некоторые из которых уже успешно использовались для решения наших задач.
В основном тексте этой рукописи мы сообщаем о 12 различных моделях. Это EfficientNets B0–B7 (Tan and Le, 2019), InceptionV3 (Szegedy et al., 2015), DenseNet121 (Huang et al., 2016), MobileNet (Sandler et al., 2018) и ResNet50 (He et al. ., 2015), обученных методом трансферного обучения (раздел 2.7). Каждая отдельная модель обучалась четыре раза с разными начальными условиями из одного и того же распределения параметров.Кроме того, мы обучили многослойные персептроны (MLP), используя в качестве входных данных 111 морфологических и цветовых признаков, упомянутых в разделе 2.4, и обучили смешанные (совместные) модели, которые объединяют MLP с более крупной моделью, обученной на изображениях (раздел 3.2). На рисунке 2 показана структура этих смешанных моделей. Наконец, мы также обучили 4-слойные сверточные сети, чтобы оценить, можем ли мы с помощью определенных видов ансамбля достичь производительности, соответствующей более крупным моделям (дополнительный раздел 5).
Рисунок 2 . Диаграмма трех основных видов моделей, которые мы упоминаем в нашей статье. Модели изображений — это сверточные сети, которые получают в качестве входных данных только изображения, модели признаков — это многослойные персептроны (MLP), которые получают в качестве входных данных только признаки, извлеченные из изображения, но не изображение как таковое , а смешанные модели объединяются и настраиваются Образные и функциональные модели.
Диаграмма трех основных видов моделей, которые мы упоминаем в нашей статье. Модели изображений — это сверточные сети, которые получают в качестве входных данных только изображения, модели признаков — это многослойные персептроны (MLP), которые получают в качестве входных данных только признаки, извлеченные из изображения, но не изображение как таковое , а смешанные модели объединяются и настраиваются Образные и функциональные модели.
2.7. Передача обучения
Поскольку обучение упомянутых моделей является очень сложной вычислительной задачей, мы использовали трансферное обучение, которое состоит из использования моделей, уже обученных распознаванию изображений в ImageNet, очень большого набора данных непланктонных изображений (Русаковский и др., 2015). Мы загрузили предварительно обученную модель и заморозили все слои. Затем мы удалили последний слой и заменили его плотным слоем с 90 463 n 90 464 90 485 90 463 c 90 464 90 488 выходами, которым предшествует и следует отсев. Новые слои (выпадение, плотность, выпадение, softmax с категориальной кросс-энтропийной потерей) и скорость обучения были оптимизированы с помощью keras-tuner (O'Malley et al., 2019). Мы запустили keras-tuner с байесовским оптимизационным поиском, 10 испытаниями и 100 эпохами, чтобы найти лучший набор гиперпараметров из байесовского поиска.Затем мы тренировались в течение 200 эпох и использовали раннюю остановку, т. е. прерывание обучения, если потери при проверке не улучшались в течение 50 эпох, и сохранение параметров модели с наименьшими потерями при проверке. Затем мы точно настроили модель, разморозив все параметры и снова переобучив ее с очень низкой скоростью обучения, η = 10 90 593 −7 90 594 , для 400 эпох.
Новые слои (выпадение, плотность, выпадение, softmax с категориальной кросс-энтропийной потерей) и скорость обучения были оптимизированы с помощью keras-tuner (O'Malley et al., 2019). Мы запустили keras-tuner с байесовским оптимизационным поиском, 10 испытаниями и 100 эпохами, чтобы найти лучший набор гиперпараметров из байесовского поиска.Затем мы тренировались в течение 200 эпох и использовали раннюю остановку, т. е. прерывание обучения, если потери при проверке не улучшались в течение 50 эпох, и сохранение параметров модели с наименьшими потерями при проверке. Затем мы точно настроили модель, разморозив все параметры и снова переобучив ее с очень низкой скоростью обучения, η = 10 90 593 −7 90 594 , для 400 эпох.
2.8. Обучение ансамблю
Ансамблевые методы используют несколько независимых алгоритмов обучения для достижения более высокой прогностической эффективности, чем можно было бы получить с помощью одного из составляющих алгоритмов обучения, что часто дает более высокие общие показатели классификации и надежность модели (Seni and Elder, 2010; Zhang and Ma, 2012). Для нашего исследования мы использовали два метода ансамбля: усреднение и суммирование.
Для нашего исследования мы использовали два метода ансамбля: усреднение и суммирование.
2.8.1. В среднем
Для каждого изображения выходом отдельной модели является n c -мерный доверительный вектор, представляющий вероятность, которую модель присваивает каждому классу. Предсказание модели является классом с наибольшей достоверностью. При усреднении ансамбля по n моделей мы берем среднее по n векторам достоверности и только после этого выбираем класс с наибольшей достоверностью.С помощью этой процедуры все модели вносят одинаковый вклад в окончательный прогноз, независимо от их производительности. Мы производили среднюю сборку на следующих вариантах моделей:
1. В разных моделях, как, например, это было успешно сделано для распознавания планктона в Lumini and Nanni (2019) и Lumini et al. (2020).
2. На разных экземплярах одной и той же модели, обученных независимо 4 раза. Это навеяно недавним наблюдением, что такой вид усреднения может привести к лучшему обобщению в моделях с достаточно большим (но не слишком большим) количеством параметров (D’Ascoli et al. , 2020). Мы обеспечиваем более глубокое обсуждение в дополнительном разделе 5.
, 2020). Мы обеспечиваем более глубокое обсуждение в дополнительном разделе 5.
3. Ручной выбор шести лучших индивидуальных моделей (на проверочном наборе) среди всех моделей. Этими лучшими моделями стали DenseNet121, EfficientNetB2, EfficientNetB5, EfficientNetB6, EfficientNetB7 и MobileNet. Для каждого мы выбрали инициализацию, обеспечивающую наилучшую производительность проверки. Мы называем это ансамблевой моделью Best_6_avg .
2.8.2. Укладка
Укладка аналогична усреднению, но каждая модель имеет разный вес.Веса определяются путем создания набора метаданных, состоящего из векторов достоверности каждой модели, и обучения полиномиальной логистической регрессии на этом наборе метаданных. Мы проводили стекирование как по начальным условиям, так и по разным архитектурам. Мы называем Best_6_stack ансамблевой моделью, полученной путем суммирования шести отдельных лучших моделей (это те же самые модели, которые мы использовали для модели Best_6_avg ).
3. Результаты
3.1. Спектакли
В таблице 1 мы обобщаем производительность отдельных моделей, а также различные формы сборки, описанные в разделе 2.8.
Таблица 1 . Точность тестирования и оценка F1 отдельных моделей в четырех различных начальных условиях.
Мы разделяем модели на три категории в зависимости от того, какие данные они принимают в качестве входных данных. Модели признаков берут числовые признаки, извлеченные из изображений, модели изображений берут обработанное изображение, а смешанные модели берут как признаки, так и изображение.
3.1.1. Производительность отдельных моделей
Во-первых, мы сосредоточимся на характеристиках одиночных моделей.Уже MLP, наша простейшая модель, которая не использует изображения в качестве входных данных, имела наилучшую точность 91,2%. Однако показатель F1 ниже 80% показывает, что точность определяется преобладающими классами.
Все модели изображений работали лучше, чем MLP, как с точки зрения точности, так и с точки зрения F1-показателя. Модель с лучшим показателем F1 — это EfficientNetB7 (F1 = 90,0%), за ней следует EfficientNetB2, которая получила почти такое же значение, но с гораздо меньшим количеством параметров (8,4 × 10 6 параметров вместо 6.6 × 10 7 параметров для EfficientNetB7). Самая легкая из представляемых нами моделей — это MobileNet с примерно 3,5 × 10 90 593 6 90 594 параметрами и максимальной оценкой F1 89,1 %.
Мы попытались еще больше повысить производительность EfficientNets, внедрив базовые методы устранения дисбаланса классов. Мы изменили вес категорий в соответствии с количеством примеров каждого класса, чтобы придать всем им равный вес, несмотря на дисбаланс классов. Мы не заметили значительных улучшений, поэтому ограничились только двумя моделями.Мы сообщаем об этом в Дополнительном разделе 4.
3.1.2. Сборка по начальным условиям
Как мы обсуждаем в дополнительном разделе 5, объединение начальных условий может помочь уменьшить разрыв в обобщении (то есть разницу между производительностью обучения и теста). Это было показано для среднего ансамбля (D'Ascoli et al., 2020; Geiger et al., 2020), но мы также проверили его для суммирования. Мы видим, что (крайние правые столбцы таблицы 1), как для суммирования, так и для усреднения, такое объединение улучшает общий результат по сравнению с производительностью каждой отдельной модели.Мы также показываем это на дополнительных рисунках 4C, D, где в каждом столбце показаны характеристики всех повторений одной модели, а также результат сборки через начальные условия. Усреднение ансамбля по (всего четырем) начальным условиям очень успешно для некоторых конкретных моделей, таких как EfficientNetB2 и DenseNet121.
Это было показано для среднего ансамбля (D'Ascoli et al., 2020; Geiger et al., 2020), но мы также проверили его для суммирования. Мы видим, что (крайние правые столбцы таблицы 1), как для суммирования, так и для усреднения, такое объединение улучшает общий результат по сравнению с производительностью каждой отдельной модели.Мы также показываем это на дополнительных рисунках 4C, D, где в каждом столбце показаны характеристики всех повторений одной модели, а также результат сборки через начальные условия. Усреднение ансамбля по (всего четырем) начальным условиям очень успешно для некоторых конкретных моделей, таких как EfficientNetB2 и DenseNet121.
3.1.3. Сборка моделей
Мы также собрали все доступные модели. Для согласованности мы сначала использовали только одно начальное условие для каждой архитектуры (выбиралось случайным образом, без повторений).Результаты, показанные в таблице 1 и дополнительных рисунках 4A, B (первые четыре столбца каждого графика), демонстрируют явное улучшение при выполнении такого рода сборки, которое в большинстве случаев кажется более эффективным, чем в начальных условиях.
3.1.4. Общая сборка
Наконец, мы объединили все модели и начальные условия, получив небольшое улучшение. Мы получили несколько лучшее улучшение при сборке шести лучших моделей из проверочного набора ( Best_6_avg и Best_6_stack ), что имело дополнительное преимущество в том, что требовало меньше ресурсов, чем при использовании всех 48 моделей.Наша последняя лучшая модель изображения, Best_6_stack , имеет точность 97,9% и показатель F1 92,7%.
С практической точки зрения производительность Best_6_avg и Best_6_stack даже лучше, чем кажется, если принять во внимание характер нашего набора данных: набор данных несбалансирован, и для большинства двух третей классов мы имеем почти идеальная классификация, как показано на рисунке 3, где мы показываем производительность по классам. Для оставшейся трети, меньшинства классов, производительность хорошая, хотя и менее надежная из-за очень небольшого количества доступных тестовых изображений. Если принять во внимание количество доступных изображений, единственные три класса с более низкой производительностью — это классы-контейнеры (или мусор): unknown, dirty, unknown_plankton. Это неудивительно, поскольку эти классы содержат большое количество различных объектов, и это не представляет большой проблемы с точки зрения мониторинга планктона, поскольку неправильные классификации, связанные с этими классами, менее актуальны (мы показываем матрицы путаницы на дополнительном рисунке). 5).
Если принять во внимание количество доступных изображений, единственные три класса с более низкой производительностью — это классы-контейнеры (или мусор): unknown, dirty, unknown_plankton. Это неудивительно, поскольку эти классы содержат большое количество различных объектов, и это не представляет большой проблемы с точки зрения мониторинга планктона, поскольку неправильные классификации, связанные с этими классами, менее актуальны (мы показываем матрицы путаницы на дополнительном рисунке). 5).
Рисунок 3 .Точность для каждого класса, отзыв и оценка F1 для моделей Best_6_avg (слева) и Best_6_stack (справа) в тестовом наборе, отсортированном на основе рисунка 1B. Поддержка, т. е. рядом с именем класса указано количество выборок истинного ответа, лежащего в каждом классе целевых значений.
Если мы исключим три класса мусора (неизвестный, грязь, неизвестный_планктон), мы получим F1-показатель = 97,3%. Если мы рассмотрим только 23 класса, для которых набор данных ZooLake содержит не менее 200 примеров (и сохраним мусорные классы с ≥200 примерами), баллы F1 возрастут до 98.0%. Наконец, если мы исключаем как классы с менее чем 200 примерами, так и мусорные классы, мы получаем F1-показатель = 98,9%.
Если мы рассмотрим только 23 класса, для которых набор данных ZooLake содержит не менее 200 примеров (и сохраним мусорные классы с ≥200 примерами), баллы F1 возрастут до 98.0%. Наконец, если мы исключаем как классы с менее чем 200 примерами, так и мусорные классы, мы получаем F1-показатель = 98,9%.
Более того, даже при ошибках наши модели не полностью отключаются. Мы можем видеть это в таблице 2, где мы отображаем первые 2 показателя моделей Best_6_avg и Best_6_stack . Они показывают, насколько хороши предположения моделей, если второй выбор классификатора считается успешным. Мы видим, что макроусредненная полнота увеличивается на 3%, а общее количество ошибочно классифицированных изображений уменьшается вдвое, при этом точность топ-2 превышает 99%.
Таблица 2 . Топ-1 и топ-2 отзыва и точности.
3.2. Смешанные модели
Поскольку наша предварительная обработка изображений не сохраняла информацию о размерах изображений, мы обучали смешанные модели, которые принимали в качестве входных данных комбинацию изображения и 111 числовых признаков, вычисленных на основе изображения.
Числовые характеристики были введены в MLP, описанный в дополнительном разделе 3, а изображения были даны в качестве входных данных для одной из моделей изображений, описанных в таблице 1.Затем две модели были объединены и объединены в плотный слой, за которым последовал софтмакс с категорической кросс-энтропийной потерей.
Используя как функции, так и изображения (без увеличения изображения) в качестве входных данных, мы тренировались с низкой скоростью обучения η = 10 90 593 −5 90 594 для 400 эпох. Для каждого выбора начальных условий каждая отдельная модель изображения была объединена с соответствующей моделью признаков (MLP). Всего мы обучили 12 смешанных моделей для 4 начальных условий каждая, всего 48 смешанных моделей.
Затем мы собрали модели и начальные условия таким же образом, как и с моделями изображений, описанными в разделе 2.8. Тестовые характеристики смешанных моделей показаны в таблице 3. Показатели одной модели немного лучше, чем результаты, полученные с помощью моделей только для изображений (таблица 1). Однако после сборки производительность смешанных моделей становится почти такой же, как у моделей изображений. Лучшая оценка F1 смешанных моделей улучшает оценку моделей изображений на 0,3%, достигая 93.0%.
Однако после сборки производительность смешанных моделей становится почти такой же, как у моделей изображений. Лучшая оценка F1 смешанных моделей улучшает оценку моделей изображений на 0,3%, достигая 93.0%.
Таблица 3 . Точность теста смешанной модели и оценка F1 отдельных моделей в четырех различных начальных условиях.
3.3. Сравнение с литературой по общедоступным наборам данных изображений морского планктона
Чтобы сравнить наш подход с предыдущей литературой, мы оценили наши модели на общедоступных наборах данных, указанных в Zheng et al. (2017), в котором сообщается о контрольных показателях классификации на ZooScan (Gorsky et al., 2010) и подмножествах Kaggle (Cowen et al., 2015) и WHOI (Sosik et al., 2014). ZooScan (Gorsky et al., 2010) состоит из 3771 изображения в градациях серого, полученных с использованием технологии Zooscan в заливе Вильфранш-сюр-Мер. Он состоит из 20 классов с переменным количеством образцов для каждого класса. Подмножество Kaggle (Zheng et al., 2017) включает 14 374 изображения в оттенках серого из 38 классов, полученных с помощью технологии In-situ Ichthyoplankton Imaging System (ISIIS) в проливе Флориды и использованных для конкурса National Data Science Bowl 2015.Распределение по классам неравномерно, но в каждом классе не менее 100 образцов. Подмножество WHOI (Sosik and Olson, 2007) содержит 6600 изображений в градациях серого разных размеров, которые были получены с помощью Imaging FlowCytobot (Olson and Sosik, 2007) из проб воды в гавани Вудс-Хоул. Подмножество содержит 22 вручную классифицированных класса планктона с одинаковым количеством образцов для каждого класса.
Подмножество Kaggle (Zheng et al., 2017) включает 14 374 изображения в оттенках серого из 38 классов, полученных с помощью технологии In-situ Ichthyoplankton Imaging System (ISIIS) в проливе Флориды и использованных для конкурса National Data Science Bowl 2015.Распределение по классам неравномерно, но в каждом классе не менее 100 образцов. Подмножество WHOI (Sosik and Olson, 2007) содержит 6600 изображений в градациях серого разных размеров, которые были получены с помощью Imaging FlowCytobot (Olson and Sosik, 2007) из проб воды в гавани Вудс-Хоул. Подмножество содержит 22 вручную классифицированных класса планктона с одинаковым количеством образцов для каждого класса.
Мы сравнили производительность наших моделей изображений с лучшими моделями Zheng et al. (2017), Lumini and Nanni (2019) и Lumini et al.(2020). Для WHOI мы использовали точно такие же наборы поездов и тестов, поскольку было доступно разделение наборов данных. Для ZooScan и Kaggle мы использовали, соответственно, двойную перекрестную проверку и пятикратную перекрестную проверку, как в Lumini et al. (2020). Мы использовали наши модели Best_6_avg и Best_6_stack и перенесли обучение, начиная с конфигураций веса, обученных на нашем наборе данных ZooLake. Мы точно настроили каждую из 6 выбранных моделей, принадлежащих Best_6_avg и Best_6_stack, со скоростью обучения η = 10 −5 , а затем усреднили и собрали стек.
(2020). Мы использовали наши модели Best_6_avg и Best_6_stack и перенесли обучение, начиная с конфигураций веса, обученных на нашем наборе данных ZooLake. Мы точно настроили каждую из 6 выбранных моделей, принадлежащих Best_6_avg и Best_6_stack, со скоростью обучения η = 10 −5 , а затем усреднили и собрали стек.
Как показано на рисунке 4, наши модели Best_6_avg и Best_6_stack всегда работали немного лучше, чем все предыдущие методы/исследования. Улучшение с точки зрения оценки F1 одинаково во всех трех наборах данных: улучшение на 1,3% по сравнению с предыдущей лучшей моделью для ZooScan, 1,0% для Kaggle и 0,3% для WHOI. Те же данные, что и на рис. 4, доступны в дополнительной таблице 2.
Рисунок 4 . Показатели Точность/оценка F1 наших моделей Best_6_avg и Best_6_stack (синие точки) в общедоступных наборах данных (ZooScan, Kaggle, WHOI) и сравнение с предыдущими результатами из литературы.Желтыми точками обозначены ансамблевые модели из (Lumini et al. , 2020): SFFS (Sequential Forward Floating Selection — метод выбора признаков, используемый для выбора моделей), WS (Weighed Selection — метод суммирования, который максимизирует производительность при минимальном количестве классификаторы). Красные точки — это модели Fus из (Lumini and Nanni, 2019), в которых сочетаются различные архитектуры и предварительная обработка. Зеленые точки обозначают нелинейное многоядерное обучение (NLMKL), где изучается оптимальная нелинейная комбинация нескольких ядер (гауссовых, полиномиальных и линейных) для объединения нескольких извлеченных признаков планктона.
, 2020): SFFS (Sequential Forward Floating Selection — метод выбора признаков, используемый для выбора моделей), WS (Weighed Selection — метод суммирования, который максимизирует производительность при минимальном количестве классификаторы). Красные точки — это модели Fus из (Lumini and Nanni, 2019), в которых сочетаются различные архитектуры и предварительная обработка. Зеленые точки обозначают нелинейное многоядерное обучение (NLMKL), где изучается оптимальная нелинейная комбинация нескольких ядер (гауссовых, полиномиальных и линейных) для объединения нескольких извлеченных признаков планктона.
Обратите внимание, что эти улучшения имеют еще одно преимущество. Наши результаты требуют объединения меньшего числа моделей и полных параметров. Средний ансамбль из 6 моделей состоял примерно из 1,58 × 10 8 параметров по сравнению с 6,25 × 10 8 (в 4,0 раза больше) лучшей модели Lumini et al. (2020) и 1,36 × 10 9 (в 8,6 раза больше) лучшей модели в Lumini and Nanni (2019). Основное преимущество более легких моделей заключается в том, что их проще развертывать и делиться ими с полевыми учеными.
Основное преимущество более легких моделей заключается в том, что их проще развертывать и делиться ими с полевыми учеными.
4. Обсуждение
В этой статье мы представили первый набор данных, насколько нам известно, изображений озерного планктона с камеры и показали, что с помощью соответствующей процедуры предварительной обработки и обучения глубоких нейронных сетей мы можем разработать модели машинного обучения, которые классифицируют их с высокой надежностью, достигая Точность 97,9% и 93,0% (макро-усредненная) оценка F1. Эти показатели улучшаются до точности 98,7% и оценки F1 96,5%, если мы исключаем несколько классов контейнеров (грязь, неизвестный, unknown_plankton), которые не идентифицируют какой-либо конкретный таксон, с оценкой F1, достигающей 98.9%, если далее ограничиться двумя третями категорий с достаточным количеством примеров.
Мы обучили несколько моделей глубокого обучения. Нашими главными нововведениями по сравнению с предыдущими приложениями к планктону являются использование моделей EfficientNet, мудрый и простой выбор модели ансамбля на этапе проверки, а также исследование методов ансамбля, вдохновленных недавней работой в области теории машинного обучения (D'Ascoli et al. ., 2020). Мы проверили полезность использования смешанных моделей, которые в качестве входных данных включают в себя, помимо изображения, числовые характеристики, такие как размер обнаруженного объекта, и обнаружили, что это увеличивает производительность одной модели, но выигрыш сглаживается, когда мы собираем в нескольких моделях (хотя лучший результат F1 все же улучшился с 92.7 до 93,0%). Мы также проверили, улучшилась ли производительность EfficientNets за счет исправления дисбаланса классов путем изменения веса классов, и не обнаружили значительного улучшения. Мы сравнили характеристики наших моделей с предыдущей литературой по наборам данных о соленой воде, получив улучшение, которое было постоянным для всех наборов данных.
., 2020). Мы проверили полезность использования смешанных моделей, которые в качестве входных данных включают в себя, помимо изображения, числовые характеристики, такие как размер обнаруженного объекта, и обнаружили, что это увеличивает производительность одной модели, но выигрыш сглаживается, когда мы собираем в нескольких моделях (хотя лучший результат F1 все же улучшился с 92.7 до 93,0%). Мы также проверили, улучшилась ли производительность EfficientNets за счет исправления дисбаланса классов путем изменения веса классов, и не обнаружили значительного улучшения. Мы сравнили характеристики наших моделей с предыдущей литературой по наборам данных о соленой воде, получив улучшение, которое было постоянным для всех наборов данных.
Наиболее эффективными отдельными моделями были EfficientNets, MobileNets и DenseNets. Примечательно, что производительность EfficientNets не зависела монотонно от количества параметров модели, возможно, из-за дисбаланса классов в нашем наборе данных. EfficientNets B2 и B7 показали лучшие результаты, но B2 использует меньшее количество параметров. Если бы нам пришлось выбирать единую архитектуру, наш выбор был бы в пользу MobileNet или EfficientNetB2, учитывая их выгодный компромисс между производительностью и размером модели. Если мы применим ансамбль, усреднение и суммирование обеспечивают аналогичные характеристики, поэтому мы предпочитаем усреднение из-за его большей простоты. Что касается смешанных моделей, их незначительное увеличение производительности после сборки, по-видимому, не оправдывает их дополнительную сложность с точки зрения развертывания.
EfficientNets B2 и B7 показали лучшие результаты, но B2 использует меньшее количество параметров. Если бы нам пришлось выбирать единую архитектуру, наш выбор был бы в пользу MobileNet или EfficientNetB2, учитывая их выгодный компромисс между производительностью и размером модели. Если мы применим ансамбль, усреднение и суммирование обеспечивают аналогичные характеристики, поэтому мы предпочитаем усреднение из-за его большей простоты. Что касается смешанных моделей, их незначительное увеличение производительности после сборки, по-видимому, не оправдывает их дополнительную сложность с точки зрения развертывания.
Системы Scripps Plankton Camera — это новая технология, которая позволяет пользователям получать большие объемы цветных изображений высокого разрешения практически с любой временной частотой. Мы заметили, что полученные нами изображения были более четкими, чем изображения, полученные в морской среде (см. Orenstein et al., 2020), что благоприятствовало процессу аннотации и классификации. Кроме того, таксономический диапазон более стабилен во время сезонной прогрессии по сравнению с морскими исследованиями: в озере присутствует меньше таксонов, чем в прибрежной морской среде, колонизация новыми таксонами относительно редка в межгодовом масштабе (новые таксоны появляются не часто), и озера того же региона разделяют большую часть состава планктонного сообщества.Это делает изучение динамики озерного планктона интересным и более контролируемым примером для разработки метода из-за его относительной экологической простоты и временной стабильности, а также подразумевает, что классификаторы озерных таксонов более надежны в этих средах в пространстве и во времени. Это особенно важно с прикладной точки зрения, поскольку инструменты, разработанные нами в этой статье, применимы не только для анализа временных рядов популяций планктона в озере Грайфензее, решения таких проблем, как вывод о взаимодействии между таксонами и прогнозирование цветения водорослей, но и могут быть переносится на другие подобные озера.
Кроме того, таксономический диапазон более стабилен во время сезонной прогрессии по сравнению с морскими исследованиями: в озере присутствует меньше таксонов, чем в прибрежной морской среде, колонизация новыми таксонами относительно редка в межгодовом масштабе (новые таксоны появляются не часто), и озера того же региона разделяют большую часть состава планктонного сообщества.Это делает изучение динамики озерного планктона интересным и более контролируемым примером для разработки метода из-за его относительной экологической простоты и временной стабильности, а также подразумевает, что классификаторы озерных таксонов более надежны в этих средах в пространстве и во времени. Это особенно важно с прикладной точки зрения, поскольку инструменты, разработанные нами в этой статье, применимы не только для анализа временных рядов популяций планктона в озере Грайфензее, решения таких проблем, как вывод о взаимодействии между таксонами и прогнозирование цветения водорослей, но и могут быть переносится на другие подобные озера. Озера представляют собой очень важные водные ресурсы для человеческого общества и требуют регулярного мониторинга качества воды и предоставления экосистемных услуг.
Озера представляют собой очень важные водные ресурсы для человеческого общества и требуют регулярного мониторинга качества воды и предоставления экосистемных услуг.
Модели, разработанные в этом исследовании, уже использовались в реальном мониторинге, для части подсчета, выполненного в Merz et al. (2021). Мы предоставляем как набор данных, так и наш код в свободном доступе.
Заявление о доступности данных
Набор данных, представленный в этом исследовании, можно найти в онлайн-репозиториях. Название репозитория и инвентарный номер можно найти ниже: https://doi.орг/10.25678/0004DY.
Вклад авторов
MB-J и FP разработали исследование. SK и MB-J построили модели для классификации зоопланктона. SK, TH, EM, TB, MR, PI, FP и MB-J активно участвовали в обсуждении при построении и улучшении моделей. TH, EM, TB, MR, PI и FP также участвовали в аннотировании изображений планктона. Все авторы внесли свой вклад в рукопись.
Финансирование
Этот проект финансировался проектом Eawag DF Big-Data Workflow (#5221. 00492.999.01), Швейцарское федеральное управление по охране окружающей среды (контракт № Q392-1149) и Швейцарский национальный научный фонд (проект 182124).
00492.999.01), Швейцарское федеральное управление по охране окружающей среды (контракт № Q392-1149) и Швейцарский национальный научный фонд (проект 182124).
Конфликт интересов
Авторы заявляют, что исследование проводилось при отсутствии каких-либо коммерческих или финансовых отношений, которые могли бы быть истолкованы как потенциальный конфликт интересов.
Примечание издателя
Все претензии, изложенные в этой статье, принадлежат исключительно авторам и не обязательно представляют претензии их дочерних организаций или издателя, редакторов и рецензентов.Любой продукт, который может быть оценен в этой статье, или претензии, которые могут быть сделаны его производителем, не гарантируются и не поддерживаются издателем.
Благодарности
Мы благодарим Т. Лоримера и С. Денниса за вклад на начальных этапах этого исследования.
Дополнительный материал
Дополнительный материал к этой статье можно найти в Интернете по адресу: https://www. frontiersin.org/articles/10.3389/fmicb.2021.746297/full#supplementary-material
frontiersin.org/articles/10.3389/fmicb.2021.746297/full#supplementary-material
Сноски
Ссылки
Абади, М., Agarwal, A., Barham, P., Brevdo, E., Chen, Z., Citro, C., et al. (2016). Tensorflow: крупномасштабное машинное обучение в гетерогенных распределенных системах. Препринт arXiv arXiv:1603.04467 . doi: 10.5281/zenodo.4724125
Полнотекстовая перекрестная ссылка | Академия Google
Банс, К. (1995). Зоопланктон: ключевая роль в контроле продукции океана: I. Биомасса и продукция. ICES J. Mar. Sci . 52, 265–277. дои: 10.1016/1054-3139(95)80043-3
Полнотекстовая перекрестная ссылка | Академия Google
Беренфельд, М.J., Randerson, J.T., McClain, C.R., Feldman, G.C., Los, S.O., Tucker, C.J., et al. (2001). Первичная продукция биосферы при энзопереходе. Наука 291, 2594–2597. doi: 10.1126/science.1055071
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Бочински Э. , Бача Г., Эйзелейн В., Уоллес Т. Дж. В., Нейстгаард Дж. К. и Сикора Т. (2019). «Глубокое активное обучение для классификации планктона in situ», в Распознавание образов и информационная криминалистика , под редакцией Z.Чжан, Д. Сутер, Ю. Тиан, А. Бранзан Албу, Н. Сидере и Х. Жаир Эскаланте (Cham. Springer International Publishing), 5–15. дои: 10.1007/978-3-030-05792-3_1
, Бача Г., Эйзелейн В., Уоллес Т. Дж. В., Нейстгаард Дж. К. и Сикора Т. (2019). «Глубокое активное обучение для классификации планктона in situ», в Распознавание образов и информационная криминалистика , под редакцией Z.Чжан, Д. Сутер, Ю. Тиан, А. Бранзан Албу, Н. Сидере и Х. Жаир Эскаланте (Cham. Springer International Publishing), 5–15. дои: 10.1007/978-3-030-05792-3_1
Полнотекстовая перекрестная ссылка | Академия Google
Cui, J., Wei, B., Wang, C., Yu, Z., Zheng, H., Zheng, B., et al. (2018). «Слияние информации о текстуре и форме сверточной нейронной сети для классификации изображений планктона», в 2018 OCEANS - MTS/IEEE Kobe Techno-Oceans (OTO) (Монтерей, Калифорния), 1–5.doi: 10.1109/OCEANSKOBE.2018.8559156
Полнотекстовая перекрестная ссылка | Академия Google
Дай Дж., Ван Р., Чжэн Х., Цзи Г. и Цяо Х. (2016). «Зоопланктонет: глубокая сверточная сеть для классификации зоопланктона», в OCEANS 2016 (Шанхай), 1–6. doi: 10.1109/OCEANSAP.2016.7485680
doi: 10.1109/OCEANSAP.2016.7485680
Полнотекстовая перекрестная ссылка | Академия Google
Дай Дж., Юй З., Чжэн Х., Чжэн Б. и Ван Н. (2017). «Гибридная сверточная нейронная сеть для классификации планктона», в Computer Vision-ACCV 2016 Workshops , под редакцией C.С. Чен, Дж. Лу и К. К. Ма (Cham: Springer International Publishing), 102–114. дои: 10.1007/978-3-319-54526-4_8
Полнотекстовая перекрестная ссылка | Академия Google
Д'Асколи С., Рефинетти М., Бироли Г. и Крзакала Ф. (2020). «Двойная проблема при двойном спуске: смещение и дисперсия (ы) в ленивом режиме», в материалах Proceedings of the 37th International Conference on Machine Learning (PMLR), 2280–2290. Доступно в Интернете по адресу: http://proceedings.mlr.press/v119/d-ascoli20a.html
.Дэвис, К.С. (1992). Видеорегистратор планктона (ВПР): конструкция и первые результаты. Арх. гидробиол. Бейх. Эргебн. Лимнол . 36, 67–81.
Дейнер, К., Бик, Х. М., Мохлер, Э., Сеймур, М., Лакурсьер-Руссель, А., Альтерматт, Ф., и другие. (2017). Экологический метабаркодирование ДНК: изменение методов исследования животных и растительных сообществ. Мол. Экол . 26, 5872–5895. doi: 10.1111/mec.14350
М., Мохлер, Э., Сеймур, М., Лакурсьер-Руссель, А., Альтерматт, Ф., и другие. (2017). Экологический метабаркодирование ДНК: изменение методов исследования животных и растительных сообществ. Мол. Экол . 26, 5872–5895. doi: 10.1111/mec.14350
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Дэн, Дж., Донг В., Сочер Р., Ли Л.-Дж., Ли К. и Фей-Фей Л. (2009). «Imagenet: крупномасштабная иерархическая база данных изображений», в 2009 IEEE Conference on Computer Vision and Pattern Recognition (Майами, Флорида: IEEE), 248–255. doi: 10.1109/CVPR.2009.5206848
Полнотекстовая перекрестная ссылка | Академия Google
Дункер, С., Бохо, Д., Вальдхен, Дж., и Мэдер, П. (2018). Сочетание высокопроизводительной проточной цитометрии изображений и глубокого обучения для эффективной идентификации видов и стадий жизненного цикла фитопланктона. БМС Экол . 18:15. doi: 10.1186/s12898-018-0209-5
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Ээрола, Т. , Крафт, К., Грёнберг, О., Ленсу, Л., Суйкканен, С., Сеппала, Дж., и др. (2020). На пути к оперативному распознаванию фитопланктона с помощью автоматизированной высокопроизводительной визуализации и компактных сверточных нейронных сетей. Науки об океане. Обсудить . 2020, 1–20. doi: 10.5194/os-2020-62
, Крафт, К., Грёнберг, О., Ленсу, Л., Суйкканен, С., Сеппала, Дж., и др. (2020). На пути к оперативному распознаванию фитопланктона с помощью автоматизированной высокопроизводительной визуализации и компактных сверточных нейронных сетей. Науки об океане. Обсудить . 2020, 1–20. doi: 10.5194/os-2020-62
Полнотекстовая перекрестная ссылка | Академия Google
Гейгер, М., Jacot, A., Spigler, S., Gabriel, F., Sagun, L., d'Ascoli, S., et al. (2020). Масштабирование описания обобщения с количеством параметров в глубоком обучении. J. Стат. мех. Теория Опыт . 2020:023401. дои: 10.1088/1742-5468/ab633c
Полнотекстовая перекрестная ссылка | Академия Google
Гонсалес П., Альварес Э., Диес Дж., Лопес-Уррутиа А. и дель Коз Дж. Дж. (2017). Методы проверки систем классификации изображений планктона. Лимнол. океаногр. Методы 15, 221–237.doi: 10.1002/lom3.10151
Полнотекстовая перекрестная ссылка | Академия Google
Гонсалес П. , Кастаньо А., Пикок Э. Э., Диез Дж., Дель Коз Дж. Дж. и Сосик Х. М. (2019). Автоматическая количественная оценка планктона с использованием глубинных признаков. Дж. Планктон Рез . 41, 449–463. doi: 10.1093/планкт/fbz023
, Кастаньо А., Пикок Э. Э., Диез Дж., Дель Коз Дж. Дж. и Сосик Х. М. (2019). Автоматическая количественная оценка планктона с использованием глубинных признаков. Дж. Планктон Рез . 41, 449–463. doi: 10.1093/планкт/fbz023
Полнотекстовая перекрестная ссылка | Академия Google
Горски Г., Оман М. Д., Пичераль М., Гаспарини С., Штемманн Л., Романьян Ж.-Б. и соавт. (2010). Анализ цифровых изображений зоопланктона с помощью интегрированной системы ZooScan. Дж. Планктон Рез . 32, 285–303. doi: 10.1093/планкт/fbp124
Полнотекстовая перекрестная ссылка | Академия Google
Го Б., Найман Л., Наяк А., Милмор Д., МакФарланд М., Твардовски М. и др. (2021). Автоматическая классификация планктона по голографическим изображениям с помощью глубоких сверточных нейронных сетей. Лимнол. океаногр. Методы 19, 21–36. doi: 10.1002/lom3.10402
Полнотекстовая перекрестная ссылка | Академия Google
Хэнд, DJ (2006). Технология классификатора и иллюзия прогресса. Стат. Наука . 21, 1–14. дои: 10.1214/088342306000000079
Стат. Наука . 21, 1–14. дои: 10.1214/088342306000000079
Полнотекстовая перекрестная ссылка | Академия Google
Хенрихс, Д. В., Англес, С., Гаонкар, К. С., и Кэмпбелл, Л. (2021). Применение сверточной нейронной сети для улучшения автоматизированного раннего предупреждения о вредоносном цветении водорослей. Окружающая среда. науч. Загрязн. Рез . 28, 28544–28555. doi: 10.1007/s11356-021-12471-2
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Хонг, С., Мехди, С.R., Huang, H., Shahani, K., Zhang, Y., Junaidullah, Raza, K., et al. (2020). Классификация пресноводного зоопланктона предварительно обученной сверточной нейронной сетью в подводной микроскопии. Междунар. Дж. Адв. вычисл. науч. Заявка . 11, 252–258. doi: 10.14569/IJACSA.2020.0110733
Полнотекстовая перекрестная ссылка | Академия Google
Хуанг Г., Лю З. и Вайнбергер К. К. (2016). Плотносвязные сверточные сети. Препринт arXiv arXiv:1608. 06993 . doi: 10.1109/CVPR.2017.243
06993 . doi: 10.1109/CVPR.2017.243
Полнотекстовая перекрестная ссылка | Академия Google
Huisman, J., Codd, G.A., Paerl, HW, Ibelings, B.W., Verspagen, JM, and Visser, P.M. (2018). Цианобактериальные цветения. Нац. Ред. Микробиол . 16, 471–483. doi: 10.1038/s41579-018-0040-1
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Керр, Т., Кларк, Дж. Р., Файлман, Э. С., Виддикомб, К. Э., и Пьюго, Н. (2020). Совместные модели глубокого обучения для устранения дисбаланса классов в изображениях планктона с потоковой камеры. Доступ IEEE 8, 170013–170032. doi: 10.1109/ACCESS.2020.3022242
Полнотекстовая перекрестная ссылка | Академия Google
Ли, Х., Парк, М., и Ким, Дж. (2016). «Классификация планктона в несбалансированной крупномасштабной базе данных с помощью сверточных нейронных сетей с передачей обучения», в 2016 IEEE International Conference on Image Processing (ICIP) (Phoenix, AZ), 3713–3717. doi: 10.1109/ICIP.2016.7533053
doi: 10.1109/ICIP.2016.7533053
Полнотекстовая перекрестная ссылка | Академия Google
Ли, Х.и Цуй, З. (2016). «Глубокие остаточные сети для классификации планктона», в OCEANS 2016 MTS/IEEE Monterey (Монтерей, Калифорния), 1–4. doi: 10.1109/OCEANS.2016.7761223
Полнотекстовая перекрестная ссылка | Академия Google
Ломбард Ф., Босс Э., Уэйт А.М., Фогт М., Уитц Дж., Штемманн Л. и др. (2019). Глобально согласованные количественные наблюдения за планктонными экосистемами. Перед. март наук . 6:196. doi: 10.3389/fmars.2019.00196
Полнотекстовая перекрестная ссылка | Академия Google
Лотце, Х.К., Титтенсор, Д.П., Бриндум-Буххольц, А., Эдди, Т.Д., Чунг, В.В., Гэлбрейт, Э.Д., и соавт. (2019). Прогнозы глобального ансамбля показывают трофическое усиление снижения биомассы океана с изменением климата. Проц. Натл. акад. науч. США . 116, 12907–12912. doi: 10.1073/pnas.1
4116Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Люмини, А. , Нанни, Л., и Магуоло, Г. (2020). Глубокое обучение для классификации планктона и кораллов. Заяв. вычисл.Сообщить . doi: 10.1016/j.aci.2019.11.004. [Epub перед печатью].
, Нанни, Л., и Магуоло, Г. (2020). Глубокое обучение для классификации планктона и кораллов. Заяв. вычисл.Сообщить . doi: 10.1016/j.aci.2019.11.004. [Epub перед печатью].
Полнотекстовая перекрестная ссылка | Академия Google
Luo, J.Y., Irisson, J.-O., Graham, B., Guigand, C., Sarafraz, A., Mader, C., et al. (2018). Автоматизированный анализ изображений планктона с использованием сверточных нейронных сетей. Лимнол. океаногр. Методы 16, 814–827. doi: 10.1002/lom3.10285
Полнотекстовая перекрестная ссылка | Академия Google
Мерц Э., Козакевич Т., Рейес М., Эби К., Айлс П., Baity-Jesi, M., et al. (2021). Подводное изображение с двойным увеличением для автоматизированного мониторинга озерного планктона. Вода Res . 203:117524. doi: 10.1016/j.waters.2021.117524
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Моншан, М.-Э., Спаак, П., и Помати, Ф. (2019). Высокие уровни рассредоточения и потепление озера являются новыми факторами формирования сообщества цианобактерий в периальпийских озерах. Науч. Реп . 9, 1–8. doi: 10.1038/s41598-019-43814-2
Науч. Реп . 9, 1–8. doi: 10.1038/s41598-019-43814-2
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Монируззаман, М., Ислам, С.М.С., Беннамун, М., и Лавери, П. (2017). «Глубокое обучение обнаружению подводных морских объектов: обзор», в Advanced Concepts for Intelligent Vision Systems , eds J. Blanc-Talon, R. Penne, W. Philips, D. Popescu, and P. Scheunders (Cham: Springer Международное издательство), 150–160. дои: 10.1007/978-3-319-70353-4_13
Полнотекстовая перекрестная ссылка | Академия Google
Морено-Торрес, Дж. Г., Редер, Т., Алаиз-Родригес, Р., Чавла, Н. В., и Эррера, Ф. (2012).Единый взгляд на изменение набора данных в классификации. Распознавание образов . 45, 521–530. doi: 10.1016/j.patcog.2011.06.019
Полнотекстовая перекрестная ссылка | Академия Google
Олсон, Р. Дж., и Сосик, Х. М. (2007). Погружной инструмент визуализации в потоке для анализа нано- и микропланктона: Imaging flowcytobot. Лимнол. океаногр. Методы 5, 195–203. doi: 10.4319/лом.2007.5.195
Лимнол. океаногр. Методы 5, 195–203. doi: 10.4319/лом.2007.5.195
Полнотекстовая перекрестная ссылка | Академия Google
О'Мэлли Т., Бурштейн Э., Long, J., Chollet, F., Jin, H., Invernizzi, L., et al. (2019). Керас Тюнер . Доступно онлайн по адресу: https://github.com/keras-team/keras-tuner
.Оренштейн, Э. К., и Бейбом, О. (2017). «Передача обучения и глубокое извлечение признаков для наборов данных планктонных изображений», в Зимняя конференция IEEE по приложениям компьютерного зрения (WACV) 2017 г. (Санта-Роза, Калифорния), 1082–1088. doi: 10.1109/WACV.2017.125
Полнотекстовая перекрестная ссылка | Академия Google
Оренштейн, Э.К., Бейбом О., Пикок Э. Э. и Сосик Х. М. (2015). Whoi-plankton: крупномасштабный мелкозернистый эталонный набор данных визуального распознавания для классификации планктона. Препринт arXiv arXiv:1510.00745 .
Orenstein, E.C., Ratelle, D., Briseno-Avena, C., Carter, M. L., Franks, P.J.S., Jaffe, J.S., et al. (2020). Система камер планктона Скриппса: основа и платформа для микроскопии in situ. Лимнол. океаногр. Методы 18, 681–695. doi: 10.1002/lom3.10394
L., Franks, P.J.S., Jaffe, J.S., et al. (2020). Система камер планктона Скриппса: основа и платформа для микроскопии in situ. Лимнол. океаногр. Методы 18, 681–695. doi: 10.1002/lom3.10394
Полнотекстовая перекрестная ссылка | Академия Google
Помати, Ф., Мэтьюз Б., Джокела Дж., Шильдкнехт А. и Ибелингс Б.В. (2012). Влияние реолиготрофирования и потепления климата на богатство планктона и стабильность сообщества в глубоком мезотрофном озере. Ойкос 121, 1317–1327. doi: 10.1111/j.1600-0706.2011.20055.x
Полнотекстовая перекрестная ссылка | Академия Google
Py, O., Hong, H., and Zhongzhi, S. (2016). «Классификация планктона с помощью глубоких сверточных нейронных сетей», 2016 IEEE Information Technology, Networking, Electronic and Automation Control Conference (Чунцин), 132–136.doi: 10.1109/ITNEC.2016.7560334
Полнотекстовая перекрестная ссылка | Академия Google
Рехт Б., Рулофс Р., Шмидт Л. и Шанкар В. (2019).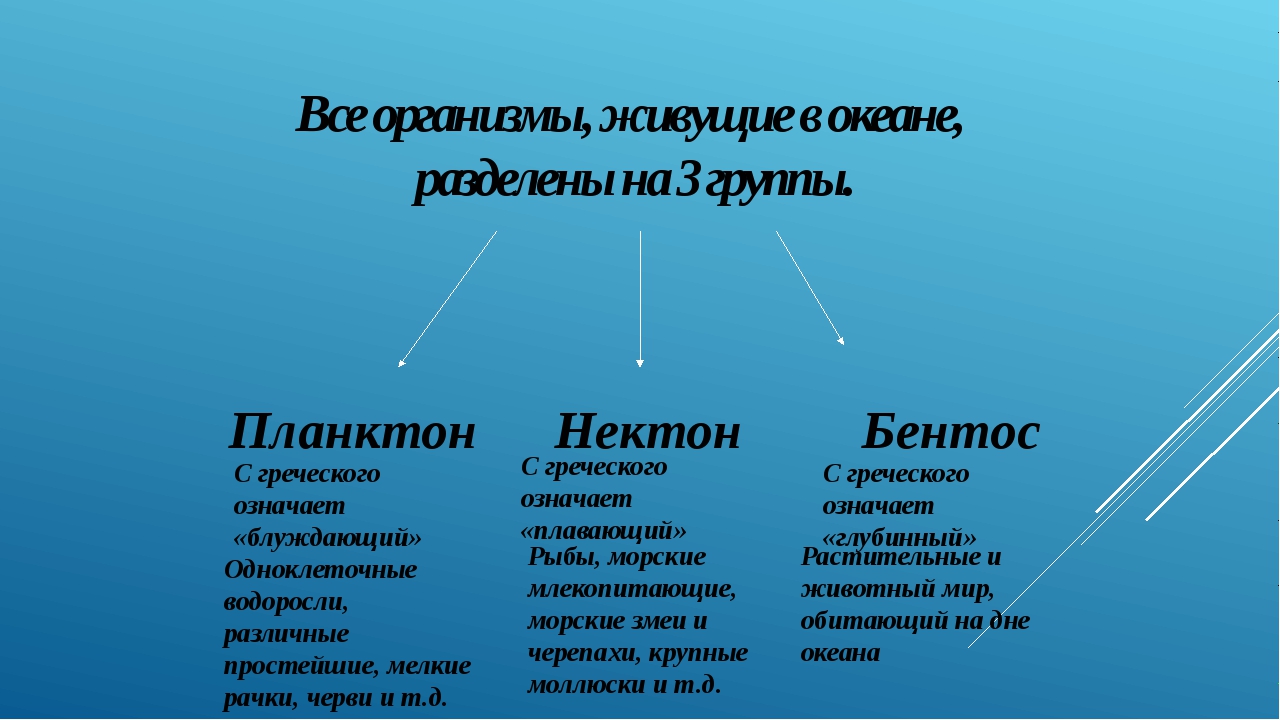 «Обобщаются ли классификаторы imagenet на imagenet?» в Международная конференция по машинному обучению (Лонг-Бич, Калифорния: PMLR), 5389–5400.
«Обобщаются ли классификаторы imagenet на imagenet?» в Международная конференция по машинному обучению (Лонг-Бич, Калифорния: PMLR), 5389–5400.
Родригес, Ф.К.М., Хирата, Н.С.Т., Абелло, А.А., Де Ла Круз, Л.Т., Лопес, Р.М., и Хирата, Р. (2018). «Оценка сценариев переноса обучения в классификации изображений планктона», в Трудах 13-й Международной объединенной конференции по теории и приложениям компьютерного зрения, обработки изображений и компьютерной графики – Том 5: VISAPP (Фуншал: INSTICC, SciTePress), 359– 366.дои: 10.5220/00066267035
Полнотекстовая перекрестная ссылка | Академия Google
Русаковский О., Денг Дж., Су Х., Краузе Дж., Сатиш С., Ма С. и др. (2015). Крупномасштабная задача визуального распознавания ImageNet. Междунар. Дж. Вычисл. Видение 115, 211–252. doi: 10.1007/s11263-015-0816-y
Полнотекстовая перекрестная ссылка | Академия Google
Салвесен, Э., Саад, А., и Шталь, А. (2020). «Надежные методы неконтролируемой кластеризации для обнаружения новых планктонных видов in situ », в Global Oceans 2020: Singapore–U. Южное побережье Мексиканского залива (Билокси, Массачусетс), 1–9. DOI: 10.1109/IEEECONF38699.2020.9389188
Южное побережье Мексиканского залива (Билокси, Массачусетс), 1–9. DOI: 10.1109/IEEECONF38699.2020.9389188
Полнотекстовая перекрестная ссылка | Академия Google
Сандлер М., Ховард А., Чжу М., Жмогинов А. и Чен Л.-К. (2018). «Mobilenetv2: перевернутые остатки и линейные узкие места», в 2018 IEEE/CVF Conference on Computer Vision and Pattern Recognition (Солт-Лейк-Сити, Юта), 4510–4520. doi: 10.1109/CVPR.2018.00474
Полнотекстовая перекрестная ссылка | Академия Google
Сосик Х.и Олсон, Р. (2007). Автоматизированная таксономическая классификация образцов фитопланктона с визуализацией «в» проточной цитометрии. Лимнол. океаногр. Методы 5, 204–216. doi: 10.4319/lom.2007.5.204
Полнотекстовая перекрестная ссылка | Академия Google
Сосик, Х.М., Пикок, Э.Э., и Браунли, Э.Ф. (2014). «Аннотированные изображения планктона — набор данных для разработки и оценки методов классификации», в WHOI-Plankton . дои: 10. 1575/1912/7341
1575/1912/7341
Полнотекстовая перекрестная ссылка | Академия Google
Сурня, А., Кретьенно-Дине, М.-Ж., и Рикар, М. (1991). Морской фитопланктон: сколько видов в мировом океане? Дж. Планктон Рез . 13, 1093–1099. doi: 10.1093/планкт/13.5.1093
Полнотекстовая перекрестная ссылка | Академия Google
Спанбауэр, Т.Л., Бриз-но-Авена, К., Питц, К.Дж., и Сутер, Э. (2020). Соленые датчики, свежие идеи: использование молекулярных датчиков и датчиков изображений для понимания динамики планктона в морских и пресноводных экосистемах. Лимнол. океаногр. Письмо .5, 169–184. дои: 10.1002/лол2.10128
Полнотекстовая перекрестная ссылка | Академия Google
Сегеди, К., Ванхуке, В., Иоффе, С., Шленс, Дж., и Война, З. (2015). Переосмысление исходной архитектуры компьютерного зрения. Препринт arXiv arXiv.1512.00567 . doi: 10.1109/CVPR.2016.308
Полнотекстовая перекрестная ссылка | Академия Google
Табак, М. А., Норуззаде, М.С., Вольфсон, Д.В., Суини, С.Дж., Веркаутерен, К.С., Сноу, Н.П., и соавт. (2019). Машинное обучение для классификации видов животных на изображениях с фотоловушек: приложения в экологии. Методы Экол. Эвол . 10, 585–590. дои: 10.1111/2041-210X.13120
А., Норуззаде, М.С., Вольфсон, Д.В., Суини, С.Дж., Веркаутерен, К.С., Сноу, Н.П., и соавт. (2019). Машинное обучение для классификации видов животных на изображениях с фотоловушек: приложения в экологии. Методы Экол. Эвол . 10, 585–590. дои: 10.1111/2041-210X.13120
Полнотекстовая перекрестная ссылка | Академия Google
Тан К., Сун Ф., Конг Т., Чжан В., Ян К. и Лю К. (2018). Опрос по глубокому трансферному обучению. Препринт arXiv arXiv.1808.01974 . дои: 10.1007/978-3-030-01424-7_27
Полнотекстовая перекрестная ссылка | Академия Google
Тан, М., и Ле, К. (2019). «EfficientNet: переосмысление масштабирования модели для сверточных нейронных сетей», в Международной конференции по машинному обучению (Лонг-Бич, Калифорния: PMLR), 6105–6114.
Тапикс, Т., Грегори-Ивз, И., и Хуот, Ю. (2021). Частная жизнь цистодиниума: наблюдение на месте за его привязанностями и динамикой популяции. Дж. Планктон Рез . 43, 492–496. doi: 10.1093/plankt/fbab025
doi: 10.1093/plankt/fbab025
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Фольк Т. и Хофферт М.И. (1985). Углеродные насосы океана: анализ относительной силы и эффективности изменений содержания CO2 в атмосфере, вызванных океаном. Углеродный цикл Атмос. CO2 физ.Вариат. Архейское настоящее 32, 99–110. дои: 10.1029/GM032p0099
Полнотекстовая перекрестная ссылка | Академия Google
Чжан, К., и Ма, Ю. (2012). Ансамблевое машинное обучение: методы и приложения . Нью-Йорк, штат Нью-Йорк: Springer-Verlag. дои: 10.1007/978-1-4419-9326-7
Полнотекстовая перекрестная ссылка | Академия Google
Чжао, Ф., Лин, Ф., и Сих, Х.С. (2010). Двоичная классификация изображений планктона сиппера с использованием случайного подпространства. Нейрокомпьютинг 73, 1853–1860.doi: 10.1016/j.neucom.2009.12.033
Полнотекстовая перекрестная ссылка | Академия Google
Чжэн, Х., Ван, Р., Ю, З.