
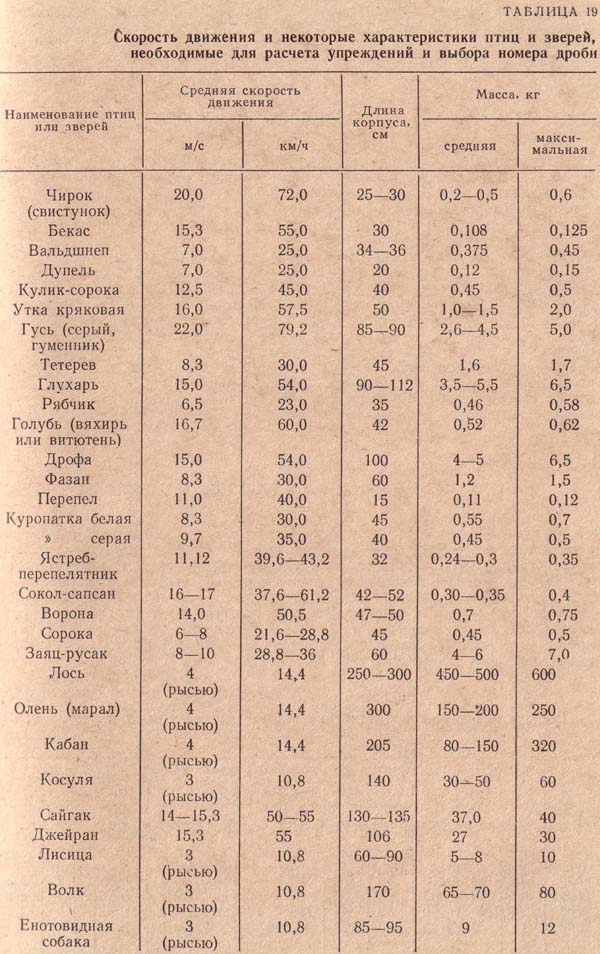
Скорость перелетаЕдва ли по какому-нибудь вопросу, связанному с перелетами птиц, распространены столь ошибочные взгляды, как по вопросу о скорости перелета. Мнения большинства людей о скорости, с которой пролетают птицы, основаны на случайных кратковременных наблюдениях, и поэтому она обычно сильно преувеличена. Другие сравнивают скорость пролета птиц со скоростью автомобиля, поезда или самолета. Однако таких скоростей они не найдут даже у наиболее быстрых из известных нам летунов. Так, например, стрижи пролетают со скоростью 40-50 м/сек (независимо от ветра), что соответствует приблизительно 150-160 км/час. При длительных миграциях имеют значение не только способность к полету, но также и ветер. В зависимости от его направления и силы скорость птиц может значительно уменьшаться или увеличиваться. Особенно большие скорости в полете можно объяснить только при учете поддержки со стороны ветра. Так, в приведенном выше примере скорость английских чибисов при полете через Атлантический океан, равная приблизительно 70 км/час, увеличилась до 150 км/час благодаря попутному ветру, скорость которого достигала 90 км/час.
Цифры, приведенные в таблице, дают ясное представление о предельных скоростях перелета птиц. Как ни мала обычно скорость перелета птиц, вернее какой малой она нам ни кажется, она вполне достаточна для того, чтобы некоторые виды за несколько дней и ночей достигли мест зимовок. Больше того, с такой скоростью при условии попутного ветра (как, например, при перелете чибисами океана) многие перелетные птицы в течение нескольких дней или ночей могли бы долететь в тропики. Однако птицы не могут сохранить указанную скорость пролета дольше, чем на несколько часов; они почти никогда не летят несколько дней или ночей подряд; как правило, их полет прерывается для короткого отдыха или для более длительных остановок; последние придают перелету в целом характер неторопливой "прогулки". При рассмотрении точно установленных кольцеванием средних скоростей дневного или ночного перелета отдельных видов всегда нужно иметь в виду, что они не характеризуют способность к полету и развиваемую на пролете скорость, а указывают лишь на длительность пролета и расстояние между местами кольцевания и находок окольцованных птиц в пересчете на один день. Многочисленные находки окольцованных птиц доказывают, что птицы быстро пролетают большую часть пути, а остальное время используют для отдыха в богатых кормом местах. Этот вид пролета встречается наиболее часто. Значительно реже наблюдается равномерное распределение нагрузки и отдыха. У птиц, летящих на большое расстояние, средний дневной путь равен примерно 150-200 км, в то время как летящие не так далеко не покрывают за это же время и 100 км. С этими данными согласуется продолжительность перелета в 2-3 или 3-4 мес. многих видов, которые зимуют в Тропической и Южной Африке. Так, например, аист, обычно отлетающий из Германии в конце августа, достигает зимовок в Южной Африке только в конце ноября или в декабре. При оценке дневного пути и скорости пролета нельзя упускать из виду и другой важный фактор: любые цифровые данные можно вычислить только для идеального пути перелета, т. е. для прямой линии, соединяющей места кольцевания и находки окольцованной птицы. В действительности пролетный путь всегда больше, отклонения от прямой часто весьма значительны и выполненная работа и скорость значительно выше вычисленных. Эти ошибки практически невозможно исключить, и поэтому их необходимо учитывать, особенно при очень длительных перелетах. Кроме того, следует обращать внимание и на то, когда были получены эти данные. Дело в том, что при весеннем перелете показатели во многих случаях значительно выше, чем при осеннем. В единичных случаях можно было бы с уверенностью доказать, что весенний перелет проходит вдвое быстрее осеннего, например у аиста, американского веретенника и жулана. Штреземан (1944) точно установил, что весной пролет жулана длится приблизительно 60 дней, а осенью - около 100 дней. В среднем эти птицы пролетают около 200 км в сутки. Однако они летят только ночью в течение 10 час. со скоростью 50 км/час. После такого пролета они всегда отдыхают, так что расстояние 1000 км покрывается ими за 5 суток: миграция - 2 ночи, сон - 3 ночи, кормежка - 5 дней. Еще несколько слов о максимальных скоростях и длительности пролета, характеризующих возможности перелетных птиц: камнешарка, небольшая прибрежная птица, окольцованная на Гельголанде, была найдена через 25 час. в Северной Франции, на 820 км южнее. Многочисленные мелкие певчие птицы регулярно перелетают за 12-15 час. Мексиканский залив шириной 750-1000 км. Согласно Моро (1938), некоторые небольшие сокола (Falco concolor и F. amurensis), а также азиатские щурки (Merops persicus и М. apiaster), зимующие на побережье Южной Африки, также пролетают не меньше 3000 км над морем. Гавайские острова служат местом, зимовки для ряда северных куликов, которые, мигрируя от Алеутских островов и Аляски, где расположены их гнездовья, вынуждены пролетать 3300 км над открытым. К пролету над водными пространствами можно приравнять пролет над большими пустынями. Такой пролег также, несомненно, проходит без перерыва, например пролет над Западной Сахарой мелких певчих птиц, трясогузок и коньков, требующий 30-40 час. непрерывной работы, если скорость их пролета считать равной приблизительно 50 км/час. |



 С этими данными согласуется продолжительность перелета в 2-3 или 3-4 мес. многих видов, которые зимуют в Тропической и Южной Африке. Так, например, аист, обычно отлетающий из Германии в конце августа, достигает зимовок в Южной Африке только в конце ноября или в декабре. Эти же сроки относятся и к жулану. Ласточки мигрируют быстрее - с сентября до начала ноября. Сколь, однако, велики в данном случае индивидуальные различия, можно видеть на примере 3 окольцованных горихвосток-лысушек, одна из которых покрывала ежедневно 167
С этими данными согласуется продолжительность перелета в 2-3 или 3-4 мес. многих видов, которые зимуют в Тропической и Южной Африке. Так, например, аист, обычно отлетающий из Германии в конце августа, достигает зимовок в Южной Африке только в конце ноября или в декабре. Эти же сроки относятся и к жулану. Ласточки мигрируют быстрее - с сентября до начала ноября. Сколь, однако, велики в данном случае индивидуальные различия, можно видеть на примере 3 окольцованных горихвосток-лысушек, одна из которых покрывала ежедневно 167  Этим данным можно противопоставить дневную скорость певчего дрозда - 40 км (вычислена за 56 дней перелета), зяблика - 17,4 км (вычислена за 23 дня перелета) и ястреба-перепелятника - 12,5 км (вычислена за 30 дней перелета). Эти данные сравнимы с приведенными выше сведениями о горихвостках, на средние скорости которых при увеличении продолжительности пролета сильно влияют длительные остановки для отдыха.
Этим данным можно противопоставить дневную скорость певчего дрозда - 40 км (вычислена за 56 дней перелета), зяблика - 17,4 км (вычислена за 23 дня перелета) и ястреба-перепелятника - 12,5 км (вычислена за 30 дней перелета). Эти данные сравнимы с приведенными выше сведениями о горихвостках, на средние скорости которых при увеличении продолжительности пролета сильно влияют длительные остановки для отдыха.
 Мексиканский залив шириной 750- 1000 км . Согласно Моро (1938), некоторые небольшие сокола (Falco concolor и F . amurensis ), а также азиатские щурки (Merops persicus и М. apiaster ), зимующие на побережье Южной Африки, также пролетают не меньше 3000 км над морем. Гавайские острова служат местом, зимовки для ряда северных куликов, которые, мигрируя от Алеутских островов и Аляски, где расположены их гнездовья, вынуждены пролетать 3300 км над открытым. морем. Золотистой ржанке, особенно сильному летуну, для покрытия этого расстояния при скорости около 90
Мексиканский залив шириной 750- 1000 км . Согласно Моро (1938), некоторые небольшие сокола (Falco concolor и F . amurensis ), а также азиатские щурки (Merops persicus и М. apiaster ), зимующие на побережье Южной Африки, также пролетают не меньше 3000 км над морем. Гавайские острова служат местом, зимовки для ряда северных куликов, которые, мигрируя от Алеутских островов и Аляски, где расположены их гнездовья, вынуждены пролетать 3300 км над открытым. морем. Золотистой ржанке, особенно сильному летуну, для покрытия этого расстояния при скорости около 90  В пути он, вероятно, совсем не отдыхает, так как в других местах его никогда не отмечали.
В пути он, вероятно, совсем не отдыхает, так как в других местах его никогда не отмечали.
 Учитывая задерживающее или ускоряющее влияние ветра, можно точно измерить собственную скорость птиц на коротких расстояниях и в соответствии с этим вычислить истинную скорость пролета. Впервые такие расчеты произвел Тинеман на Курской косе. Впоследствии они были сделаны Мейнертцхагеном, Гаррисоном и др
Учитывая задерживающее или ускоряющее влияние ветра, можно точно измерить собственную скорость птиц на коротких расстояниях и в соответствии с этим вычислить истинную скорость пролета. Впервые такие расчеты произвел Тинеман на Курской косе. Впоследствии они были сделаны Мейнертцхагеном, Гаррисоном и др В общем она, очевидно, равна 40-80 км/час, причем скорость мелких певчих птиц приближается к низшим цифрам. Птицы, совершающие перелет ночью, летят, по-видимому, быстрее, чем мигрирующие днем. Бросается в глаза малая скорость миграции хищных и других крупных птиц. Одни и те же виды птиц летают в области гнездования обычно значительно медленнее, чем на пролете, если вообще эти скорости можно сравнивать.
В общем она, очевидно, равна 40-80 км/час, причем скорость мелких певчих птиц приближается к низшим цифрам. Птицы, совершающие перелет ночью, летят, по-видимому, быстрее, чем мигрирующие днем. Бросается в глаза малая скорость миграции хищных и других крупных птиц. Одни и те же виды птиц летают в области гнездования обычно значительно медленнее, чем на пролете, если вообще эти скорости можно сравнивать. Так возникают длительные миграции.
Так возникают длительные миграции. Эти же сроки относятся и к жулану. Ласточки мигрируют быстрее - с сентября до начала ноября. Сколь, однако, велики в данном случае индивидуальные различия, можно видеть на примере 3 окольцованных горихвосток-лысушек, одна из которых покрывала ежедневно 167 км, другая - 61 км и третья - лишь 44 км, причем эти числа уменьшаются по мере увеличения отрезка времени, для которого их вычисляют (6, 30 и 47 дней). Основываясь на этих результатах, можно сделать вывод, что дневная скорость более всего соответствует истинной скорости перелета в том случае, когда она вычисляется на основе общих показателей за короткий срок. Этот вывод лучше всего доказывают следующие примеры скорости пролетов отдельных птиц: аист покрыл за 2 дня 610 км, славка-черноголовка за 10 дней - 2200 км, лысуха за 7 дней - 1300 км, другая лысуха за 2 дня - 525 км, кряква за 5 дней - 1600 км. Этим данным можно противопоставить дневную скорость певчего дрозда - 40 км (вычислена за 56 дней перелета), зяблика - 17,4 км (вычислена за 23 дня перелета) и ястреба-перепелятника - 12,5 км (вычислена за 30 дней перелета).
Эти же сроки относятся и к жулану. Ласточки мигрируют быстрее - с сентября до начала ноября. Сколь, однако, велики в данном случае индивидуальные различия, можно видеть на примере 3 окольцованных горихвосток-лысушек, одна из которых покрывала ежедневно 167 км, другая - 61 км и третья - лишь 44 км, причем эти числа уменьшаются по мере увеличения отрезка времени, для которого их вычисляют (6, 30 и 47 дней). Основываясь на этих результатах, можно сделать вывод, что дневная скорость более всего соответствует истинной скорости перелета в том случае, когда она вычисляется на основе общих показателей за короткий срок. Этот вывод лучше всего доказывают следующие примеры скорости пролетов отдельных птиц: аист покрыл за 2 дня 610 км, славка-черноголовка за 10 дней - 2200 км, лысуха за 7 дней - 1300 км, другая лысуха за 2 дня - 525 км, кряква за 5 дней - 1600 км. Этим данным можно противопоставить дневную скорость певчего дрозда - 40 км (вычислена за 56 дней перелета), зяблика - 17,4 км (вычислена за 23 дня перелета) и ястреба-перепелятника - 12,5 км (вычислена за 30 дней перелета). Эти данные сравнимы с приведенными выше сведениями о горихвостках, на средние скорости которых при увеличении продолжительности пролета сильно влияют длительные остановки для отдыха.
Эти данные сравнимы с приведенными выше сведениями о горихвостках, на средние скорости которых при увеличении продолжительности пролета сильно влияют длительные остановки для отдыха.
 морем. Золотистой ржанке, особенно сильному летуну, для покрытия этого расстояния при скорости около 90 км/час потребовалось бы приблизительно 35 час. Более высокие скорости отмечены у другого вида ржанок, пролетающих от Новой Шотландии до северной оконечности Южной Америки 3600 км над морем. Почти невероятным кажется пролет одного из гнездящихся в Японии бекасов, который зимует в Восточной Австралии и должен покрыть почти 5000 км, чтобы достигнуть зимовок. В пути он, вероятно, совсем не отдыхает, так как в других местах его никогда не отмечали.
морем. Золотистой ржанке, особенно сильному летуну, для покрытия этого расстояния при скорости около 90 км/час потребовалось бы приблизительно 35 час. Более высокие скорости отмечены у другого вида ржанок, пролетающих от Новой Шотландии до северной оконечности Южной Америки 3600 км над морем. Почти невероятным кажется пролет одного из гнездящихся в Японии бекасов, который зимует в Восточной Австралии и должен покрыть почти 5000 км, чтобы достигнуть зимовок. В пути он, вероятно, совсем не отдыхает, так как в других местах его никогда не отмечали. Кольцеванием и наблюдениями удалось во многих случаях точно доказать наличие регулярного, хотя и не очень интенсивного пролета над Альпами, Кавказом и даже над мощными горными цепями Гималаев. Следует особенно подчеркнуть, что птицы далеко не всегда предпочитают долины и перевалы, а пролетают и над горными хребтами (например, в Тянь-Шане), на высоте 6000 м . В сравнении с ними высоты, преодолеваемые в Альпах, кажутся незначительными (Наибольшей высоты 9500 м , на которую когда-либо поднимались птицы, достигла стая гусей над Эверестом. Высоту удалось точно определить, стая гусей была сфотографирована с самолета (Гаррисон, 1931).).
Кольцеванием и наблюдениями удалось во многих случаях точно доказать наличие регулярного, хотя и не очень интенсивного пролета над Альпами, Кавказом и даже над мощными горными цепями Гималаев. Следует особенно подчеркнуть, что птицы далеко не всегда предпочитают долины и перевалы, а пролетают и над горными хребтами (например, в Тянь-Шане), на высоте 6000 м . В сравнении с ними высоты, преодолеваемые в Альпах, кажутся незначительными (Наибольшей высоты 9500 м , на которую когда-либо поднимались птицы, достигла стая гусей над Эверестом. Высоту удалось точно определить, стая гусей была сфотографирована с самолета (Гаррисон, 1931).). Во всяком случае, абсолютная высота сама по себе как будто не оказывает на высоту пролета птиц существенного влияния. Одни и те же виды птиц летят на равнине на такой же высоте, как и в горах, если условия ветра и погоды существенно не различаются.
Во всяком случае, абсолютная высота сама по себе как будто не оказывает на высоту пролета птиц существенного влияния. Одни и те же виды птиц летят на равнине на такой же высоте, как и в горах, если условия ветра и погоды существенно не различаются. Для перепелятников эти высоты составляли соответственно 640 и 850 м , а такие крупные птицы, как канюк и бородач, на высоте 1500-2000 м были еле заметны.
Для перепелятников эти высоты составляли соответственно 640 и 850 м , а такие крупные птицы, как канюк и бородач, на высоте 1500-2000 м были еле заметны. При помощи современных авиационных измерительных приборов во время второй мировой войны часто удавалось установить перелеты птиц на больших высотах, например вяхиря на высоте 2500 м . Подводя итог, можно, однако, утверждать, что пролеты птиц на высоте более 1000 м сравнительно редки. В норме пролеты проходят на высоте нескольких сот метров, а часто, особенно у мелких птиц, ниже 100 м . При благоприятной погоде и хорошей видимости и не слишком сильном ветре птицы летят значительно выше, чем при низкой облачности, дожде, тумане или более сильном встречном ветре. Чем сильнее ветер, тем ниже летят птицы, используя каждую возвышенность, опушки лесов и долины рек, где сила ветра несколько слабее. Над морем птицы также летят большей частью над поверхностью воды. Они по возможности избегают туманов и полетов в облаках. Над сплошным слоем облаков редко можно встретить пролетных птиц, но иногда над густой облачностью летят крупные птицы, например гуси и журавли.
При помощи современных авиационных измерительных приборов во время второй мировой войны часто удавалось установить перелеты птиц на больших высотах, например вяхиря на высоте 2500 м . Подводя итог, можно, однако, утверждать, что пролеты птиц на высоте более 1000 м сравнительно редки. В норме пролеты проходят на высоте нескольких сот метров, а часто, особенно у мелких птиц, ниже 100 м . При благоприятной погоде и хорошей видимости и не слишком сильном ветре птицы летят значительно выше, чем при низкой облачности, дожде, тумане или более сильном встречном ветре. Чем сильнее ветер, тем ниже летят птицы, используя каждую возвышенность, опушки лесов и долины рек, где сила ветра несколько слабее. Над морем птицы также летят большей частью над поверхностью воды. Они по возможности избегают туманов и полетов в облаках. Над сплошным слоем облаков редко можно встретить пролетных птиц, но иногда над густой облачностью летят крупные птицы, например гуси и журавли. Триб указывал на улучшение видимости на дальние расстояния при закате солнца, а также на хорошую видимость красных сигнальных огней в туманную погоду. Это явление объясняется тем, что длинноволновые желтые и красные лучи лучше проникают через туманную атмосферу, чем коротковолновые зеленые, синие и фиолетовые. При этом безразлично, рассматривается ли местность при красноватом свете или же так, чтобы желтые и красные лучи оказывались особенно действенными. Так, например, поступает фотограф, который при съемке ландшафта с затуманенной далью экспонирует светочувствительную пластинку через оранжевый светофильтр. Если исключить все лучи, кроме наиболее длинных в инфракрасной части спектра, то можно фотографировать даже на расстоянии сотен километров. Подобное действие оказывают и красно-желтые шарики в глазах птиц. Таким образом, перелетные птицы могут даже при туманной погоде из Сицилии видеть берега Африки. Тем не менее создается впечатление, что птицы ориентируются главным образом по особым признакам ландшафта, а не по общим очертаниям поверхности земли.
Триб указывал на улучшение видимости на дальние расстояния при закате солнца, а также на хорошую видимость красных сигнальных огней в туманную погоду. Это явление объясняется тем, что длинноволновые желтые и красные лучи лучше проникают через туманную атмосферу, чем коротковолновые зеленые, синие и фиолетовые. При этом безразлично, рассматривается ли местность при красноватом свете или же так, чтобы желтые и красные лучи оказывались особенно действенными. Так, например, поступает фотограф, который при съемке ландшафта с затуманенной далью экспонирует светочувствительную пластинку через оранжевый светофильтр. Если исключить все лучи, кроме наиболее длинных в инфракрасной части спектра, то можно фотографировать даже на расстоянии сотен километров. Подобное действие оказывают и красно-желтые шарики в глазах птиц. Таким образом, перелетные птицы могут даже при туманной погоде из Сицилии видеть берега Африки. Тем не менее создается впечатление, что птицы ориентируются главным образом по особым признакам ландшафта, а не по общим очертаниям поверхности земли. Если бы это было иначе, то некоторые острова не играли бы роли направляющих линий и направляющих пунктов, исчезли бы массовые пролетные пути, а многочисленные обходные пути сократились бы. Однако все указанные условия существуют и оказывают влияние на перелеты птиц, являясь доказательством того, что перелеты проходят в среднем на небольшой высоте. Часто птицам достаточно подняться лишь немного выше, чтобы найти ориентиры, которые, казалось бы, делают излишним следование по направляющим линиям. Дункер (1905) составил сводку дальности видимости (вне зависимости от атмосферных условий) с различных высот. На основании формулы r = КОРЕНЬ 2Rk = 113 КОРЕНЬ h получены следующие цифры.
Если бы это было иначе, то некоторые острова не играли бы роли направляющих линий и направляющих пунктов, исчезли бы массовые пролетные пути, а многочисленные обходные пути сократились бы. Однако все указанные условия существуют и оказывают влияние на перелеты птиц, являясь доказательством того, что перелеты проходят в среднем на небольшой высоте. Часто птицам достаточно подняться лишь немного выше, чтобы найти ориентиры, которые, казалось бы, делают излишним следование по направляющим линиям. Дункер (1905) составил сводку дальности видимости (вне зависимости от атмосферных условий) с различных высот. На основании формулы r = КОРЕНЬ 2Rk = 113 КОРЕНЬ h получены следующие цифры. Город для них стал уже родным домом, это не только источник пищи, но и источник гнездования. На чердаках домов регулярно все заселенно голубями, им там тепло и хорошо. Гнезда зачастую построены далеко не из природных материалов, а из мелкого мусора, ниток, полиэтилена, строительного мусора, утеплены ватой и стекловатой, торчащей из утепленных окон или поломанной обшивки домов. Эти птички, так же как и другие гости (трясогузки, снегири, свиристели, поползни и др.), которых можно встретить в городской среде, обитают не небольших высотах - возвышаясь до 10-16 этажей (до 50 метров). Выше им просто уже нет смысла взлетать, там нет не еды, ни гнезд.
Город для них стал уже родным домом, это не только источник пищи, но и источник гнездования. На чердаках домов регулярно все заселенно голубями, им там тепло и хорошо. Гнезда зачастую построены далеко не из природных материалов, а из мелкого мусора, ниток, полиэтилена, строительного мусора, утеплены ватой и стекловатой, торчащей из утепленных окон или поломанной обшивки домов. Эти птички, так же как и другие гости (трясогузки, снегири, свиристели, поползни и др.), которых можно встретить в городской среде, обитают не небольших высотах - возвышаясь до 10-16 этажей (до 50 метров). Выше им просто уже нет смысла взлетать, там нет не еды, ни гнезд.| Синицы на кормушке |
Голуби, вороны, ласточки и коршуны
Тоже частые обитатели городов. Если голубей мы все время видим под ногами, то ворон уже преимущественно на деревьях, а коршунов только в небесах. Птицы по крупнее способны перелетать на более крупные расстояния, и чтоб им легче летелось, взмывают выше. Здесь уже речь идет о высоте в 70-100 метров и выше. Для сравнения самое высокое здание в Новосибирске - Коммунистическая 50 (Бэтмен) имеет высоты 87,9 метров.
Здесь уже речь идет о высоте в 70-100 метров и выше. Для сравнения самое высокое здание в Новосибирске - Коммунистическая 50 (Бэтмен) имеет высоты 87,9 метров.
| Коршун, парящий в небесах |
Самые высоко летающие птицы
Существуют птицы, которые взлетают еще выше! В каждодневной жизни они поднимаются на высоты 1000-1500 метров, а для длительных перелетов еще выше. Выглядят они совершенно по другому - они крупные, у них большой размах крыльев, все это позволяет им накапливать много энергии, летать далеко, высоко и долго.
- Белый аист поднимается на высоту 2-3 тыс. метров.
- Черные стрижи - рекордсмены среди птиц по длительности пребывания в воздухе (молодые стрижи, взлетев, приземляются только через 2-3 года), вторые по скорости полета (больше 120 км/ч) и одни из самых высоко летающих птиц мира (могут кружить даже во сне на высоте около 3 тысяч метров над землей).

- Беркут - это хищная птица, которая кружа на высоте 4500 метров, может своим острым зрением заметить добычу и ее поймать.
- Кряквы - многим известные и встречаемые даже в пределах города утки. Во время перелетов летают очень высоко. Известен случай, когда кряква столкнулась с самолетом на высоте 6900 метров.
- Серые гуси перелетая поднимаются на высоту 8000 метров.
- Самые рекордсмены это грифы, максимальная высота на которой они были зафиксированы 12150 метров, столкнувшись с самолетом.

Гражданские самолеты же летают на высоте 9-11 тыс. км. Во время перелета, каждый кто летал, смотрел или хотя бы заглядывал в окошко, разглядеть внизу что-то очень трудно. Город виден красивым только при взлете и посадке, когда самолет постепенно набирает высоту или ее снижает.
Так каких птиц имеют в виду люди, когда вводят запрос в Google? Про какую высоту они говорят? 🙂
С детства нам интересна одна простая загадка: кто же на самом деле самый быстрый из птиц? Эти удивительные существа имеют такой запас сил, что многие могут им позавидовать. Результат исследований на подобную тему может многих удивить.
Результат исследований на подобную тему может многих удивить.
Самая быстрая птица в мире
Первое место в подобном списке самых быстрых птиц занимает сапсан. Именно эта неприметная птичка может перемещаться со скоростью приблизительно 389 километров в час, что (для сравнения) значительно превышает скорость свободного падения парашютистов.
Именно эта самая быстрая птица в мире может дать фору многим животным, при этом сапсана можно встретить абсолютно на всех континентах, кроме Антарктиды разве что. Главной его особенностью является то, что он может развивать такую огромную скорость, только пикируя с высоты.
Размеры сапсана
На вид эта самая быстрая птица в мире не больше вороны, к тому же она имеет серое оперение, которое на брюшке становится светло-серым, а голова всегда черного цвета.
Выживает сапсан благодаря своеобразной технике охоты, которая заключается в том, что он пикирует с высоты на свою жертву и сбивает ее с ног ударом поджатых лапок. Скорость, с которой сапсан это делает, может запросто привести к тому, что он собьет голову бедной добыче.
Скорость, с которой сапсан это делает, может запросто привести к тому, что он собьет голову бедной добыче.
Второй по скорости
На самом деле, птица, о которой пойдет речь дальше, может спокойно занять и первое место этого своеобразного рейтинга скорости.
Главной причиной этого является то, что сапсан развивает огромную скорость, когда «падает» с небес, а вот скорость полета стрижа огромна в горизонтальной плоскости.
Он может развивать скорость более 170 километров в час. Встретиться с таким чудом можно только в Северной или же Средней Азии, а также в Средней Европе. Свое зимовье птица проводит в Африке или же в Индии. Теперь уже естественным ареалом ее обитания являются города, и намного реже это касается лесов.
Внешний вид стрижа
Стриж имеет размер еще меньше, чем сапсан, и весит всего лишь 50-150 грамм.
Черный стриж является самым быстрым. Он имеет оперение темно-бурого оттенка с еле заметным металлическим отливом. Его легко можно спутать с ласточкой, ведь эти виды птиц достаточно похожи, особенно если смотреть сверху.
Особенности птицы
Особенность стрижа именно в том, что буквально несколько веков назад его могли употреблять в пищу, аргументируя тем, что мясо достаточно вкусное.
Если не учитывать этой его гастрономической особенности, есть и другой любопытный факт: практически все свое время стриж проводит в воздухе. В самом буквальном смысле этого слова. Вылетая через восемь недель после рождения из гнезда, он приземляется приблизительно только через 3 года. С условием того, что у него лапки очень короткие и пальцы направлены только вперед, самостоятельно взлететь с земли достаточно трудно, но можно. Необходимо лишь несколько очень сильных взмахов крыльями и хотя бы небольшая возвышенность для того, чтобы легче было взлететь. Крылья сами по себе непропорционально большие, если все-таки их сравнивать с размерами самого тела.
Длинные, изогнутой формы крылья и идеально обтекаемое тело, плоская голова, а также короткая шея - все эти аэродинамические особенности позволяют стрижу даже спать в воздухе. Находясь на высоте до 3 тысяч метров в стайке, он просто летает по кругу и засыпает, при этом просыпается каждые 5 секунд для того, чтобы в очередной раз взмахнуть крыльями и не упасть.
Находясь на высоте до 3 тысяч метров в стайке, он просто летает по кругу и засыпает, при этом просыпается каждые 5 секунд для того, чтобы в очередной раз взмахнуть крыльями и не упасть.
Эта самая быстрая птица на земле может за всю свою жизнь пролететь приблизительно 500 тысяч километров, при этом приземлившись всего лишь несколько раз в жизни только для того, чтобы вывести потомство.
Ласточка и стриж: отличия
Ранее говорилось о том, что стриж может быть очень сильно похож на ласточку своим внешним видом. Но все-таки самым важным их отличием является скорость полета - стриж развивает скорость около 170 километров в час, а ласточка всего лишь 60 километров в час. Но тем не менее, стриж не может быть лучше по маневренности полета, чем ласточка. Птица этого вида также отличается от стрижа строением лапок - у стрижа четыре пальца повернуты вперед, а у ласточек - три вперед и один назад. Именно поэтому они могут садиться на телеграфные провода и там легко удерживаться, а вот у стрижа подобное точно не получится.
У стрижей темное брюшко, а у ласточек оно белое. При этом в полете первый вид птичек отличается своей излишней крикливостью и никогда не складывает крылья. Кроме того, стриж больше по размерам, чем ласточка.
Скорость полета птиц
Третьим по скорости в этом рейтинге является сероголовый альбатрос. Он больше по размерам, чем его предшественники, ведь имеет размах крыльев в 3,5 метра. Из-за того что альбатрос не может совершать такого стремительного пикирования либо же постоянно жить в воздухе, он отличается своей выносливостью.
Именно он может летать со скоростью в 130 километров в час на протяжении восьми часов. Он хоть и не самая быстрая птица в мире, но состоит в Книге рекордов Гиннеса в связи со своими удивительными особенностями.
Гага - это птица из семейства утиных, которая способна развивать скорость приблизительно до ста километров в час. При этом она выдерживает длительные перелеты, хотя и не поднимается высоко в небо, ведь ее основная пища находится в воде - моллюски, черви, мелкие рыбешки. Именно поэтому гага - не только быстрая птица, но еще и отличный ныряльщик.
Именно поэтому гага - не только быстрая птица, но еще и отличный ныряльщик.
Следующим в рейтинге самых быстрых птиц мира будет почтовый голубь. Этот вид отлично себя зарекомендовал в различных условиях - и в мирное время, и во время военных действий. Именно поэтому голубю нужно отдать должное уважение.
Скорость его полета составляет от 90 и до 100 километров в час. Голуби намного выносливее, чем альбатросы, - отдельные особи могут находиться в воздухе более 16 часов.
Скворец - это незаметная птичка с приятным певчим голосом, она также зарекомендовала себя в качестве отличного летуна. Скворцы могут развивать скорость приблизительно в 70 километров в час, и встречаются они на каждом континенте нашей планеты.
Также скорость в 70 километров в час может развивать и дрозд-рябинник. Он распространен на всей территории Европы и Азии, а его голос и необычное оперение всегда привлекают достаточно много внимания.
Скорость перелетаЕдва ли по какому-нибудь вопросу, связанному с перелетами птиц, распространены столь ошибочные взгляды, как по вопросу о скорости перелета. Мнения большинства людей о скорости, с которой пролетают птицы, основаны на случайных кратковременных наблюдениях, и поэтому она обычно сильно преувеличена. Другие сравнивают скорость пролета птиц со скоростью автомобиля, поезда или самолета. Однако таких скоростей они не найдут даже у наиболее быстрых из известных нам летунов. Так, например, стрижи пролетают со скоростью 40-50 м/сек (независимо от ветра), что соответствует приблизительно 150-160 км/час. (Сравните: максимальная скорость экспресса - 39 м/сек, или 140 км/час. При длительных миграциях имеют значение не только способность к полету, но также и ветер. В зависимости от его направления и силы скорость птиц может значительно уменьшаться или увеличиваться. Особенно большие скорости в полете можно объяснить только при учете поддержки со стороны ветра. Так, в приведенном выше примере скорость английских чибисов при полете через Атлантический океан, равная приблизительно 70 км/час, увеличилась до 150 км/час благодаря попутному ветру, скорость которого достигала 90 км/час. Учитывая задерживающее или ускоряющее влияние ветра, можно точно измерить собственную скорость птиц на коротких расстояниях и в соответствии с этим вычислить истинную скорость пролета.
Цифры, приведенные в таблице, дают ясное представление о предельных скоростях перелета птиц. Как ни мала обычно скорость перелета птиц, вернее какой малой она нам ни кажется, она вполне достаточна для того, чтобы некоторые виды за несколько дней и ночей достигли мест зимовок. Больше того, с такой скоростью при условии попутного ветра (как, например, при перелете чибисами океана) многие перелетные птицы в течение нескольких дней или ночей могли бы долететь в тропики. Однако птицы не могут сохранить указанную скорость пролета дольше, чем на несколько часов; они почти никогда не летят несколько дней или ночей подряд; как правило, их полет прерывается для короткого отдыха или для более длительных остановок; последние придают перелету в целом характер неторопливой "прогулки". При рассмотрении точно установленных кольцеванием средних скоростей дневного или ночного перелета отдельных видов всегда нужно иметь в виду, что они не характеризуют способность к полету и развиваемую на пролете скорость, а указывают лишь на длительность пролета и расстояние между местами кольцевания и находок окольцованных птиц в пересчете на один день. Многочисленные находки окольцованных птиц доказывают, что птицы быстро пролетают большую часть пути, а остальное время используют для отдыха в богатых кормом местах. Этот вид пролета встречается наиболее часто. Значительно реже наблюдается равномерное распределение нагрузки и отдыха. У птиц, летящих на большое расстояние, средний дневной путь равен примерно 150-200 км, в то время как летящие не так далеко не покрывают за это же время и 100 км. С этими данными согласуется продолжительность перелета в 2-3 или 3-4 мес. многих видов, которые зимуют в Тропической и Южной Африке. Так, например, аист, обычно отлетающий из Германии в конце августа, достигает зимовок в Южной Африке только в конце ноября или в декабре. При оценке дневного пути и скорости пролета нельзя упускать из виду и другой важный фактор: любые цифровые данные можно вычислить только для идеального пути перелета, т. е. для прямой линии, соединяющей места кольцевания и находки окольцованной птицы. В действительности пролетный путь всегда больше, отклонения от прямой часто весьма значительны и выполненная работа и скорость значительно выше вычисленных. Эти ошибки практически невозможно исключить, и поэтому их необходимо учитывать, особенно при очень длительных перелетах. Кроме того, следует обращать внимание и на то, когда были получены эти данные. Дело в том, что при весеннем перелете показатели во многих случаях значительно выше, чем при осеннем. В единичных случаях можно было бы с уверенностью доказать, что весенний перелет проходит вдвое быстрее осеннего, например у аиста, американского веретенника и жулана. Штреземан (1944) точно установил, что весной пролет жулана длится приблизительно 60 дней, а осенью - около 100 дней. В среднем эти птицы пролетают около 200 км в сутки. Однако они летят только ночью в течение 10 час. со скоростью 50 км/час. После такого пролета они всегда отдыхают, так что расстояние 1000 км покрывается ими за 5 суток: миграция - 2 ночи, сон - 3 ночи, кормежка - 5 дней. Еще несколько слов о максимальных скоростях и длительности пролета, характеризующих возможности перелетных птиц: камнешарка, небольшая прибрежная птица, окольцованная на Гельголанде, была найдена через 25 час. в Северной Франции, на 820 км южнее. Многочисленные мелкие певчие птицы регулярно перелетают за 12-15 час. Мексиканский залив шириной 750-1000 км. Согласно Моро (1938), некоторые небольшие сокола (Falco concolor и F. amurensis), а также азиатские щурки (Merops persicus и М. apiaster), зимующие на побережье Южной Африки, также пролетают не меньше 3000 км над морем. Гавайские острова служат местом, зимовки для ряда северных куликов, которые, мигрируя от Алеутских островов и Аляски, где расположены их гнездовья, вынуждены пролетать 3300 км над открытым. К пролету над водными пространствами можно приравнять пролет над большими пустынями. Такой пролег также, несомненно, проходит без перерыва, например пролет над Западной Сахарой мелких певчих птиц, трясогузок и коньков, требующий 30-40 час. непрерывной работы, если скорость их пролета считать равной приблизительно 50 км/час. |
 На нее очень сильно влияют атмосферные явления, поэтому при дальних перемещениях птицы то летят быстрее, то медленнее, то делают длительные перерывы для отдыха.
На нее очень сильно влияют атмосферные явления, поэтому при дальних перемещениях птицы то летят быстрее, то медленнее, то делают длительные перерывы для отдыха. Так же как и скорость, ее очень трудно измерить. Сокол, выпущенный под Парижем, через день был обнаружен на острове Мальта за 1400 км. Задерживался он в пути или летел все время, неизвестно. Вообще птицы останавливаются в пути довольно часто, и отрезки беспосадочных перелетов у них невелики. Этого нельзя сказать о перелетах через водные преграды, где птицам негде сесть. Рекорд на дальность беспосадочного перелета принадлежит куликам - бурокрылым ржанкам, ежегодно пролетающим над океаном с Аляски на Гавайи и обратно 3000 км. Птицы перелетают без посадки через Мексиканский залив (1300 км), Средиземное море (600-750 км), Северное море (600 км), Черное море (300 км). Значит, средняя дальность беспосадочного перелета птиц составляет около 1000 км.
Так же как и скорость, ее очень трудно измерить. Сокол, выпущенный под Парижем, через день был обнаружен на острове Мальта за 1400 км. Задерживался он в пути или летел все время, неизвестно. Вообще птицы останавливаются в пути довольно часто, и отрезки беспосадочных перелетов у них невелики. Этого нельзя сказать о перелетах через водные преграды, где птицам негде сесть. Рекорд на дальность беспосадочного перелета принадлежит куликам - бурокрылым ржанкам, ежегодно пролетающим над океаном с Аляски на Гавайи и обратно 3000 км. Птицы перелетают без посадки через Мексиканский залив (1300 км), Средиземное море (600-750 км), Северное море (600 км), Черное море (300 км). Значит, средняя дальность беспосадочного перелета птиц составляет около 1000 км. Гриф вывел из строя один из моторов, но самолет благополучно приземлился. Это, видимо, абсолютный рекорд высоты полета птиц. До этого бородач был отмечен в Гималаях на высоте 7900 м, пролетные гуси там же на высоте 9500 м, кряква столкнулась с самолетом над Невадой на высоте 6900 м.
Гриф вывел из строя один из моторов, но самолет благополучно приземлился. Это, видимо, абсолютный рекорд высоты полета птиц. До этого бородач был отмечен в Гималаях на высоте 7900 м, пролетные гуси там же на высоте 9500 м, кряква столкнулась с самолетом над Невадой на высоте 6900 м.
 Теперь уже естественным ареалом ее обитания являются города, и намного реже это касается лесов.
Теперь уже естественным ареалом ее обитания являются города, и намного реже это касается лесов. Необходимо лишь несколько очень сильных взмахов крыльями и хотя бы небольшая возвышенность для того, чтобы легче было взлететь. Крылья сами по себе непропорционально большие, если все-таки их сравнивать с размерами самого тела.
Необходимо лишь несколько очень сильных взмахов крыльями и хотя бы небольшая возвышенность для того, чтобы легче было взлететь. Крылья сами по себе непропорционально большие, если все-таки их сравнивать с размерами самого тела. Но тем не менее, стриж не может быть лучше по маневренности полета, чем ласточка. Птица этого вида также отличается от стрижа строением лапок - у стрижа четыре пальца повернуты вперед, а у ласточек - три вперед и один назад. Именно поэтому они могут садиться на телеграфные провода и там легко удерживаться, а вот у стрижа подобное точно не получится.
Но тем не менее, стриж не может быть лучше по маневренности полета, чем ласточка. Птица этого вида также отличается от стрижа строением лапок - у стрижа четыре пальца повернуты вперед, а у ласточек - три вперед и один назад. Именно поэтому они могут садиться на телеграфные провода и там легко удерживаться, а вот у стрижа подобное точно не получится. Он хоть и не самая быстрая птица в мире, но состоит в Книге рекордов Гиннеса в связи со своими удивительными особенностями.
Он хоть и не самая быстрая птица в мире, но состоит в Книге рекордов Гиннеса в связи со своими удивительными особенностями. Скворцы могут развивать скорость приблизительно в 70 километров в час, и встречаются они на каждом континенте нашей планеты.
Скворцы могут развивать скорость приблизительно в 70 километров в час, и встречаются они на каждом континенте нашей планеты. ) Это, конечно, не означает, что птицы вообще не умеют летать быстрее. Гоняющиеся друг за другом стрижи развивают скорость до 200 км/час, а сокол бросается на жертву со скоростью 70 м/сек, т. е. 250 км/час. Но эти предельные скорости в течение очень короткого времени являются исключениями: они в лучшем случае характеризуют способность к полету некоторых видов, но их нельзя использовать для оценки скорости полета при миграциях, когда необходимо длительное напряжение.
) Это, конечно, не означает, что птицы вообще не умеют летать быстрее. Гоняющиеся друг за другом стрижи развивают скорость до 200 км/час, а сокол бросается на жертву со скоростью 70 м/сек, т. е. 250 км/час. Но эти предельные скорости в течение очень короткого времени являются исключениями: они в лучшем случае характеризуют способность к полету некоторых видов, но их нельзя использовать для оценки скорости полета при миграциях, когда необходимо длительное напряжение. Впервые такие расчеты произвел Тинеман на Курской косе. Впоследствии они были сделаны Мейнертцхагеном, Гаррисоном и др
Впервые такие расчеты произвел Тинеман на Курской косе. Впоследствии они были сделаны Мейнертцхагеном, Гаррисоном и др В общем она, очевидно, равна 40-80 км/час, причем скорость мелких певчих птиц приближается к низшим цифрам. Птицы, совершающие перелет ночью, летят, по-видимому, быстрее, чем мигрирующие днем. Бросается в глаза малая скорость миграции хищных и других крупных птиц. Одни и те же виды птиц летают в области гнездования обычно значительно медленнее, чем на пролете, если вообще эти скорости можно сравнивать.
В общем она, очевидно, равна 40-80 км/час, причем скорость мелких певчих птиц приближается к низшим цифрам. Птицы, совершающие перелет ночью, летят, по-видимому, быстрее, чем мигрирующие днем. Бросается в глаза малая скорость миграции хищных и других крупных птиц. Одни и те же виды птиц летают в области гнездования обычно значительно медленнее, чем на пролете, если вообще эти скорости можно сравнивать. Так возникают длительные миграции.
Так возникают длительные миграции. Эти же сроки относятся и к жулану. Ласточки мигрируют быстрее - с сентября до начала ноября. Сколь, однако, велики в данном случае индивидуальные различия, можно видеть на примере 3 окольцованных горихвосток-лысушек, одна из которых покрывала ежедневно 167 км, другая - 61 км и третья - лишь 44 км, причем эти числа уменьшаются по мере увеличения отрезка времени, для которого их вычисляют (6, 30 и 47 дней). Основываясь на этих результатах, можно сделать вывод, что дневная скорость более всего соответствует истинной скорости перелета в том случае, когда она вычисляется на основе общих показателей за короткий срок. Этот вывод лучше всего доказывают следующие примеры скорости пролетов отдельных птиц: аист покрыл за 2 дня 610 км, славка-черноголовка за 10 дней - 2200 км, лысуха за 7 дней - 1300 км, другая лысуха за 2 дня - 525 км, кряква за 5 дней - 1600 км. Этим данным можно противопоставить дневную скорость певчего дрозда - 40 км (вычислена за 56 дней перелета), зяблика - 17,4 км (вычислена за 23 дня перелета) и ястреба-перепелятника - 12,5 км (вычислена за 30 дней перелета).
Эти же сроки относятся и к жулану. Ласточки мигрируют быстрее - с сентября до начала ноября. Сколь, однако, велики в данном случае индивидуальные различия, можно видеть на примере 3 окольцованных горихвосток-лысушек, одна из которых покрывала ежедневно 167 км, другая - 61 км и третья - лишь 44 км, причем эти числа уменьшаются по мере увеличения отрезка времени, для которого их вычисляют (6, 30 и 47 дней). Основываясь на этих результатах, можно сделать вывод, что дневная скорость более всего соответствует истинной скорости перелета в том случае, когда она вычисляется на основе общих показателей за короткий срок. Этот вывод лучше всего доказывают следующие примеры скорости пролетов отдельных птиц: аист покрыл за 2 дня 610 км, славка-черноголовка за 10 дней - 2200 км, лысуха за 7 дней - 1300 км, другая лысуха за 2 дня - 525 км, кряква за 5 дней - 1600 км. Этим данным можно противопоставить дневную скорость певчего дрозда - 40 км (вычислена за 56 дней перелета), зяблика - 17,4 км (вычислена за 23 дня перелета) и ястреба-перепелятника - 12,5 км (вычислена за 30 дней перелета). Эти данные сравнимы с приведенными выше сведениями о горихвостках, на средние скорости которых при увеличении продолжительности пролета сильно влияют длительные остановки для отдыха.
Эти данные сравнимы с приведенными выше сведениями о горихвостках, на средние скорости которых при увеличении продолжительности пролета сильно влияют длительные остановки для отдыха.
 морем. Золотистой ржанке, особенно сильному летуну, для покрытия этого расстояния при скорости около 90 км/час потребовалось бы приблизительно 35 час. Более высокие скорости отмечены у другого вида ржанок, пролетающих от Новой Шотландии до северной оконечности Южной Америки 3600 км над морем. Почти невероятным кажется пролет одного из гнездящихся в Японии бекасов, который зимует в Восточной Австралии и должен покрыть почти 5000 км, чтобы достигнуть зимовок. В пути он, вероятно, совсем не отдыхает, так как в других местах его никогда не отмечали.
морем. Золотистой ржанке, особенно сильному летуну, для покрытия этого расстояния при скорости около 90 км/час потребовалось бы приблизительно 35 час. Более высокие скорости отмечены у другого вида ржанок, пролетающих от Новой Шотландии до северной оконечности Южной Америки 3600 км над морем. Почти невероятным кажется пролет одного из гнездящихся в Японии бекасов, который зимует в Восточной Австралии и должен покрыть почти 5000 км, чтобы достигнуть зимовок. В пути он, вероятно, совсем не отдыхает, так как в других местах его никогда не отмечали. Очень важно, чтобы ученики поняли, что поддержание постоянной температуры тела обеспечивается взаимодействием целого ряда физиологических систем организма. Хорошее знание этого материала необходимо для объяснения сложных эволюционных и экологических проблем.
Очень важно, чтобы ученики поняли, что поддержание постоянной температуры тела обеспечивается взаимодействием целого ряда физиологических систем организма. Хорошее знание этого материала необходимо для объяснения сложных эволюционных и экологических проблем. )
)
 Насекомоядные птицы наполняют свой желудок 5–6 раз в сутки, зерноядные – трижды.
Насекомоядные птицы наполняют свой желудок 5–6 раз в сутки, зерноядные – трижды. 5).
5). Вот несколько примеров.
Вот несколько примеров.  С его помощью сова по писку и шороху узнает о присутствии грызунов и точно определяет место нахождения добычи.
С его помощью сова по писку и шороху узнает о присутствии грызунов и точно определяет место нахождения добычи. Затем отнес яйцо к большой скале и швырнул об нее! Когда и это не принесло желаемого результата, стервятник принялся отчаянно колотить одно яйцо о другое.
Затем отнес яйцо к большой скале и швырнул об нее! Когда и это не принесло желаемого результата, стервятник принялся отчаянно колотить одно яйцо о другое.

 Этого нельзя сказать о перелетах через водные преграды, где птицам негде сесть. Рекорд на дальность беспосадочного перелета принадлежит куликам — бурокрылым ржанкам, ежегодно пролетающим над океаном с Аляски на Гавайи и обратно 3000 км.
Этого нельзя сказать о перелетах через водные преграды, где птицам негде сесть. Рекорд на дальность беспосадочного перелета принадлежит куликам — бурокрылым ржанкам, ежегодно пролетающим над океаном с Аляски на Гавайи и обратно 3000 км.







 Эти ошибки практически невозможно исключить, и поэтому их необходимо учитывать, особенно при очень длительных перелетах.
Эти ошибки практически невозможно исключить, и поэтому их необходимо учитывать, особенно при очень длительных перелетах.


 — Під час залицяння птахи здійснюють акробатичні трюки у польоті, граються зі здобиччю».
— Під час залицяння птахи здійснюють акробатичні трюки у польоті, граються зі здобиччю».



 Чаще всего откладывают 2-4 красно-коричневых яиц с красными точками.
Чаще всего откладывают 2-4 красно-коричневых яиц с красными точками. В погоне за добычей сапсан в момент пикирования может достичь огромных скоростей! Максимальная зарегистрированная скорость сокола-сапсана в пике составляет 389 км/ч!
В погоне за добычей сапсан в момент пикирования может достичь огромных скоростей! Максимальная зарегистрированная скорость сокола-сапсана в пике составляет 389 км/ч! Свою жертву сапсан может распознать даже с высоты 300 метров.
Свою жертву сапсан может распознать даже с высоты 300 метров.
 Пустельга летает, чередуя медленный или быстрый полет со скольжением. Иногда птицы могут парить и даже зависать в воздухе, тогда хвост как бы подвисает, а крылья совершают быстрые мелкие взмахи («вибрируют»). Обыкновенная пустельга может также двигаться по инерции, сложив крылья, или лететь при неполном их раскрытии.
Пустельга летает, чередуя медленный или быстрый полет со скольжением. Иногда птицы могут парить и даже зависать в воздухе, тогда хвост как бы подвисает, а крылья совершают быстрые мелкие взмахи («вибрируют»). Обыкновенная пустельга может также двигаться по инерции, сложив крылья, или лететь при неполном их раскрытии. Скорость этих птиц около 70 км/ч, при этом они несколько больше скворцов по размерам (25 см) и размаху крыльев (39-42 см).
Скорость этих птиц около 70 км/ч, при этом они несколько больше скворцов по размерам (25 см) и размаху крыльев (39-42 см). Они способны проводить в небе до 16 часов. Интересно, что почтовым голубям воздвигнуто больше памятников, чем другим пернатым. Мало того, памятник почтовому голубю есть почти в каждом европейском городе. Большая их часть связана с заслугами почтовых голубей во время Второй мировой войны.
Они способны проводить в небе до 16 часов. Интересно, что почтовым голубям воздвигнуто больше памятников, чем другим пернатым. Мало того, памятник почтовому голубю есть почти в каждом европейском городе. Большая их часть связана с заслугами почтовых голубей во время Второй мировой войны. Этот факт был установлен благодаря датчикам слежения, прикрепленным к лапе одной из птиц этого вида. Эти данные даже зафиксированы в Книге рекордов Гиннеса.
Этот факт был установлен благодаря датчикам слежения, прикрепленным к лапе одной из птиц этого вида. Эти данные даже зафиксированы в Книге рекордов Гиннеса. Удар он наносит сложенными и прижатыми к туловищу лапами. По точным вычислениям, падая на добычу под углом в 25°, сокол летит со скоростью 75 метров в секунду; при падении под углом, близким к прямому, скорость увеличивается до 100 метров в секунду или 360 км/ч. Есть данные, что сапсан способен развить скорость до 440 км/ч, что сравнимо со скоростью некоторых самолётов. Сапсаны начинают преследование добычи уже с расстояния от одного до полутора километров.
Удар он наносит сложенными и прижатыми к туловищу лапами. По точным вычислениям, падая на добычу под углом в 25°, сокол летит со скоростью 75 метров в секунду; при падении под углом, близким к прямому, скорость увеличивается до 100 метров в секунду или 360 км/ч. Есть данные, что сапсан способен развить скорость до 440 км/ч, что сравнимо со скоростью некоторых самолётов. Сапсаны начинают преследование добычи уже с расстояния от одного до полутора километров.Высота полета птиц таблица
Ежедневно в разных уголках нашей планеты в небо поднимаются сотни тысяч, а то и миллионы птиц. Крылья этих существ позволяют им не только преодолевать огромные расстояния, но и подниматься вверх на большие высоты. Их уникальные способности в навигации до сих пор поражают многих ученых, а порой остаются для них все такой же загадкой.
Мы уже привыкли наблюдать за высоко парящими в небе птицами. Но как высоко они могут подниматься в небо, и какие именно птицы поднимаются выше остальных пернатых? В большинстве случаев они летят на высоте около 150 метров, но вовремя ежегодных миграций, некоторые представители способны подняться на высоту и до 3 тысяч метров.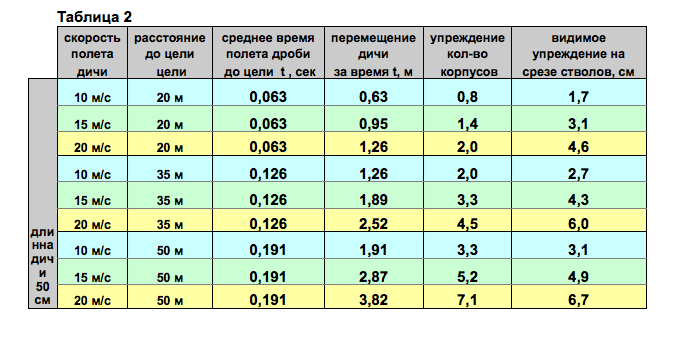 Но как оказалось и это не предел…
Но как оказалось и это не предел…
Белый аист
Белый аист (лат. Ciconia ciconia) – одна из самых красивых и грациозных птиц планеты. Этот длинношеий и длинноногий представитель пернатого мира во время дальних перелетов к местам зимовки преодолевает огромнейшие расстояния на высоте от 2 до 3 тысяч метров. Сразу хочется отметить, что размах крыльев белого аиста составляет 150-200 см.
Андский кондор
Андский кондор (лат. Vultur gryphus) – одна из самых крупных летающих хищных птиц Западного полушария. Кроме этого андский кондор является национальным символом таких латиноамериканских государств как Аргентина, Боливия, Чили, Колумбия, Эквадор и Перу, играя важнейшую роль в культуре всех этих стран. Эта уникальная птица, живя в более чем суровых местах Анд, ежедневно в поисках пищи совершает перелеты по несколько километров, подымаясь на высоту около 3-5 тысяч метров.
Черный стриж
Черные стрижи (лат. Apus apus) – эту небольшую птицу еще называют «бешеным стрижем» и это не удивительно, поскольку черный стриж считается рекордсменом среди пернатых по длительности пребывания в воздухе и вторым по скорости полета (более 120 км/час). Кроме того черный стриж – одна из самых высоко летающих птиц. Он поднимается в высоту до 3000 метров.
Кроме того черный стриж – одна из самых высоко летающих птиц. Он поднимается в высоту до 3000 метров.
Беркут
Беркут (лат. Aquila chrysaetos) – еще одна крупная и наиболее известная хищная птица, с способная подниматься на большие высоты. Размах крыльев беркута может достигать двух метров. У этого хищника прекрасное зрение, поэтому ему не составляет труда подняться на высоту 4500 метров над землей и оттуда высматривать свою добычу.
Ржанки
Ржанки (лат. Pluvialis) – небольшая, но симпатичная птица из семейства ржанковых (лат. Charadriidae). Может летать как на совершенно низком расстоянии (едва не касаясь поверхности воды), так и на высоте более 6 тысяч метров. Представители этого семейства хорошо известны на ваттовых побережьях (Ваттовом море).
Кряква
Кряква (лат. Anas platyrhynchos) – птица принадлежащая семейству утиных (лат. Anatidae) и самая узнаваемая и распространенная дикая утка.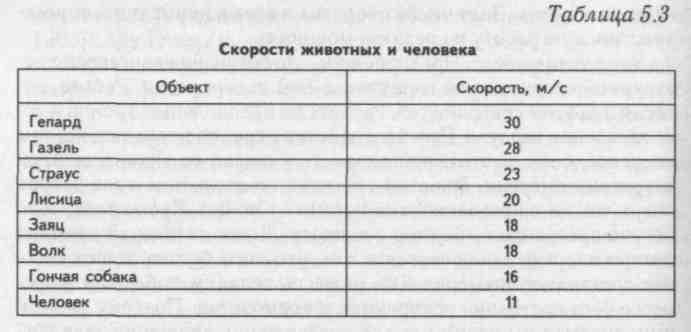 Однако мало кто знает, что эта водоплавающая птица также является прекрасным летуном. Во время ежегодных перелетов на места зимовки эти птицы поднимаются в высоту до 6900 метров. Бывали случаи, когда утки на такой высоте сталкивались с летящим самолетом.
Однако мало кто знает, что эта водоплавающая птица также является прекрасным летуном. Во время ежегодных перелетов на места зимовки эти птицы поднимаются в высоту до 6900 метров. Бывали случаи, когда утки на такой высоте сталкивались с летящим самолетом.
Серые гуси
Серые гуси (лат. Anser anser) – представитель из числа водоплавающих птиц. Селятся эти гуси по берегам озер, болт, прудов и других водоемов. Это очень умные, сильные и осторожные птицы. Они легко могут дать отпор даже хищнику, особенно если тот угрожает их потомству. Во время миграций серые гуси, собравшись в стаи, летят клином, врассыпную или шеренгой. Во время такого перелета птицы поднимаются на высоту до 8 тысяч метров.
Лебедь-кликун
Лебедь-кликун (лат. Cygnus cygnus) – крупная (от 7 до 10 кг) водоплавающая птица. В декабре 1967 года была замечена небольшая стая лебедей, пролетавшая на высоте 8230 метров над Ирландией. Высоту полета птиц зафиксировали радары. Стоит отметить, что это был не единственный зафиксированный полет лебедей-крикунов, и их способность летать на высоте более 8 тысяч метров была подтверждена не раз.
Стоит отметить, что это был не единственный зафиксированный полет лебедей-крикунов, и их способность летать на высоте более 8 тысяч метров была подтверждена не раз.
Горный гусь
Горный гусь (лат. Eulabeia indica) – водоплавающая птица, гнездящаяся в горах Центральной Азии (на высоте от 1000 до 5000 м). По словам ученых, эта птица способна пролететь над Гималаями всего за 8 часов, поднимаясь на высоту 10175 (!) метров. В то же время, они не способны преодолевать очень большие расстояния.
Гриф Рюппеля, или африканский сип
Гриф Рюппеля, или африканский сип (лат. Gyps rueppellii) – самые-самые высоко летающие птицы планеты. По утверждению ученых, именно грифы Рюппеля чаще всего сталкиваются с летящими самолетами. Самые большие зафиксированные высоты полета этого грифа – 11277 метров и 12150 метров. Обитает африканский сип в северных и восточных частях Африканского континента.
Такие полеты поистине вызывают восторг. Но как птицам, летящим на таких высотах, удается справляться с разряженным воздухом, солнечным излучением и низкими температурами пока неизвестно.
Скорость, дальность, высота полета птиц
Относительно скорости полета птиц исследователи придерживаются различных мнений. На нее очень сильно влияют атмосферные явления, поэтому при дальних перемещениях птицы то летят быстрее, то медленнее, то делают длительные перерывы для отдыха.
Выпустив птицу в каком-то месте, очень трудно сказать, когда она прилетит в "пункт назначения", ведь она может лететь далеко не все время своего отсутствия. Скорость, вычисленная путем простого деления расстояния на время перелета птицы, часто бывает заниженной. В особенно "ответственные" моменты — преследуя добычу или спасаясь от опасности — птицы могут развивать и очень большие скорости, но, конечно, долго их не выдерживают. Крупные соколы во время ставки — преследования птицы в воздухе — достигают скоростей в 280-360 км/ч. Обычные, "повседневные" скорости птиц средней величины гораздо меньше — 50-90 км/ч.
Все сказанное выше касалось машущего полета. Скорость скользящего полета также трудно поддается измерению. Считают, что чеглок планирует со скоростью 150 км/ч, бородач-ягнятник — 140, а гриф — даже 250 км/ч.
Считают, что чеглок планирует со скоростью 150 км/ч, бородач-ягнятник — 140, а гриф — даже 250 км/ч.
Дальность беспосадочных перелетов птиц обсуждается уже давно. Так же как и скорость, ее очень трудно измерить. Сокол, выпущенный под Парижем, через день был обнаружен на острове Мальта за 1400 км. Задерживался он в пути или летел все время, неизвестно. Вообще птицы останавливаются в пути довольно часто, и отрезки беспосадочных перелетов у них невелики. Этого нельзя сказать о перелетах через водные преграды, где птицам негде сесть. Рекорд на дальность беспосадочного перелета принадлежит куликам — бурокрылым ржанкам, ежегодно пролетающим над океаном с Аляски на Гавайи и обратно 3000 км. Птицы перелетают без посадки через Мексиканский залив (1300 км), Средиземное море (600-750 км), Северное море (600 км), Черное море (300 км). Значит, средняя дальность беспосадочного перелета птиц составляет около 1000 км.
Как правило, высота полета птиц не достигает 1000 м. Но отдельные крупные хищники, гуси, утки могут подниматься и на значительно большие высоты. В сентябре 1973 г. африканский гриф столкнулся с гражданским самолетом на высоте 12 150 м над Берегом Слоновой Кости. Гриф вывел из строя один из моторов, но самолет благополучно приземлился. Это, видимо, абсолютный рекорд высоты полета птиц. До этого бородач был отмечен в Гималаях на высоте 7900 м, пролетные гуси там же на высоте 9500 м, кряква столкнулась с самолетом над Невадой на высоте 6900 м.
В сентябре 1973 г. африканский гриф столкнулся с гражданским самолетом на высоте 12 150 м над Берегом Слоновой Кости. Гриф вывел из строя один из моторов, но самолет благополучно приземлился. Это, видимо, абсолютный рекорд высоты полета птиц. До этого бородач был отмечен в Гималаях на высоте 7900 м, пролетные гуси там же на высоте 9500 м, кряква столкнулась с самолетом над Невадой на высоте 6900 м.
Человек всегда стремился покорять мир, в котором он живёт: углубиться в недры земли, погрузиться на дно океана, покорить самую высокую гору. И лишь одно осталось недоступным для человечества — это полёт. Умение парить на большой высоте без специального оборудования и аппаратуры дано только птицам. Кто они, эти птицы высокого полета и как далеко они могут подняться расскажем в списке самых высоко летающих птиц мира
Белый аист – десятое место
Белый аист — это типичный обитатель умеренных широт. Особенностью этих птиц является своеобразный тип миграции, а именно, очень быстрый перелёт от мест гнездования к местам отдыха в Восточной Африке. Это около 4,5 тысяч км, которое птицы преодолевают за 18-19 дней, находясь ежедневно в пути по 8-10 часов. Длительные остановки, более суток, делаются только при плохих погодных условиях.
Это около 4,5 тысяч км, которое птицы преодолевают за 18-19 дней, находясь ежедневно в пути по 8-10 часов. Длительные остановки, более суток, делаются только при плохих погодных условиях.
Андский кондор – девятое место
Кондор по праву считается гигантом, так как по весу и размеру превосходит всех птиц Западного Полушария. Птица является обитателем горной гряды Анд. Селится преимущественно на открытых плоскогорьях.
Кондор относится к падальщикам, и пользуется уважением среди птиц этого типа питания благодаря своему мощному клюву, зачастую приступает к трапезе первым. Ест долго, так как его желудок вмещает до 4-х кг пищи за один раз.
Малый веретенник – восьмое место
Ареал обитания этих птиц очень велик. Они гнездятся в тундре и лесотундре, на территории от Скандинавского полуострова и до Аляски, в некоторых районах Европейской части России. Местом их зимовки стал берег Средиземного моря, атлантическое побережье Африки, Красное море, побережье Индии, Австралии и Новой Зеландии.
Эти птицы являются рекордсменами по непрерывному преодолению больших расстояний. Длина пути миграции этих птиц превышает 11000 км, а скорость ее преодоления всего 7-8 дней.
Кряква – седьмое место
Самый узнаваемый тип дикой утки гнездится в разных климатических зонах: как в арктических широтах, так и в субтропиках. Несмотря на свою неуклюжесть на суше, в воздухе способна преодолевать большие расстояния. Известны даже случаи столкновения уток с самолетом. Особенностью полета птиц является характерный звук, издаваемый крыльями уток, так еще не видя птиц в воздухе охотники вычисляют их приближение.
Бородач – шестое место
Эта редкая птица является уникальной в роде ястребиных. Относится к исчезающему виду. Внимание биологов привлек метод питания этих хищников.
Серые гуси – пятое место
Размеры: 70-90 см
Вес: 2,1-4,5 кг
Размах крыльев: 147-180 см
Максимальная высота полета: до 8000 м.
Средой обитания гусей являются водоемы со стоячей водой – болота, озера, водохранилища. Обычный полет серого гуся существенно отличается от сезонных перелетов. Так летая низко с редким взмахом крыла, во время миграции птицы поднимаются на большую высоту и парят в воздухе. Несмотря на внешнюю безобидность, отчаянно защищают свое потомство. Способны нанести ущерб небольшому хищнику или охотничьей собаке.
Обычный полет серого гуся существенно отличается от сезонных перелетов. Так летая низко с редким взмахом крыла, во время миграции птицы поднимаются на большую высоту и парят в воздухе. Несмотря на внешнюю безобидность, отчаянно защищают свое потомство. Способны нанести ущерб небольшому хищнику или охотничьей собаке.
Лебедь-кликун – четвертое место
Этот вид лебедей достаточно распространен – обитает в средней части Евразии и Европы. Местами миграции являются побережья Черного, Красного, Средиземного море, а также Атлантического и Тихого океана.
Свое название птица получила за громкие трубные звуки, обычно издаваемые в полёте. Неторопливо плавающий лебедь, при приближении опасности способен развивать большую скорость. Не каждая лодка способна догнать испуганную птицу.
Горный гусь – третье место
Этот вид птиц населяет горные водоемы центральной Азии. Горный гусь может преодолеть расстояние в 1600 км при низком атмосферном давлении за один день. Стоит отметить, что горный воздух настолько разрежен, что вертолет не летает на такие высоты, однако эти птицы способны в зимние время перелетать горный массив Гималаев.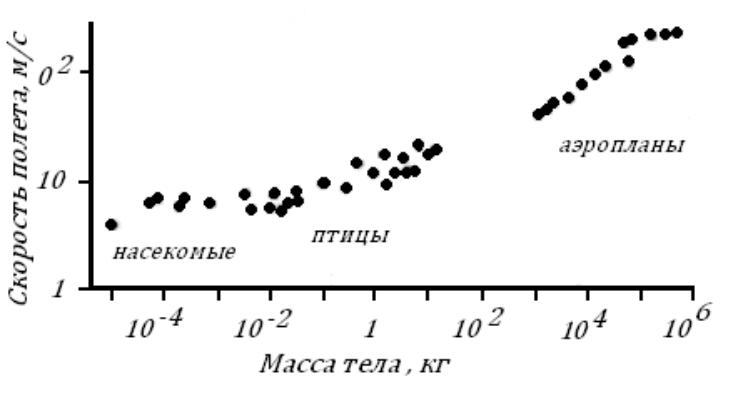
Серый журавль – второе место
Места гнездования журавлей замечены в Северной и Западной Европе, также почти на всей на территории России и Китая, и местами в Турции, на Алтае и Тибете. Мигрирует через Гималаи на юго-восток Китая, на побережье Тихого океана. Сегодня является исчезающим видом по причине сокращения территории обитания.
Самая высоко летающая птица в мире
Самая высоко летающая птица в мире это африканский сип. Местом обитания птицы является Центральная Африка. Организм птицы способен выдерживать экстремально низкий уровень кислорода. Причиной тому является особый вид гемоглобина, содержащийся в крови грифа. Установлено, что подъем на такую высоту необходим птице для лучшего обзора территории и своевременного обнаружения добычи.
Самые высоко летающие птицы
Птицы были и остаются пока что единственными хозяевами неба. В ТОП 3 входят горный гусь, серый журавль и африканский сип. Ученым остается догадываться, как без аэродинамических чертежей и сложных математических подсчетов, природе удалось создать таких уникальных в своем роде существ.
Скорость перелетаЕдва ли по какому-нибудь вопросу, связанному с перелетами птиц, распространены столь ошибочные взгляды, как по вопросу о скорости перелета. Мнения большинства людей о скорости, с которой пролетают птицы, основаны на случайных кратковременных наблюдениях, и поэтому она обычно сильно преувеличена. Другие сравнивают скорость пролета птиц со скоростью автомобиля, поезда или самолета. Однако таких скоростей они не найдут даже у наиболее быстрых из известных нам летунов. Так, например, стрижи пролетают со скоростью 40-50 м/сек (независимо от ветра), что соответствует приблизительно 150-160 км/час. При длительных миграциях имеют значение не только способность к полету, но также и ветер. В зависимости от его направления и силы скорость птиц может значительно уменьшаться или увеличиваться. Особенно большие скорости в полете можно объяснить только при учете поддержки со стороны ветра. Так, в приведенном выше примере скорость английских чибисов при полете через Атлантический океан, равная приблизительно 70 км/час, увеличилась до 150 км/час благодаря попутному ветру, скорость которого достигала 90 км/час.
Цифры, приведенные в таблице, дают ясное представление о предельных скоростях перелета птиц. Как ни мала обычно скорость перелета птиц, вернее какой малой она нам ни кажется, она вполне достаточна для того, чтобы некоторые виды за несколько дней и ночей достигли мест зимовок. Больше того, с такой скоростью при условии попутного ветра (как, например, при перелете чибисами океана) многие перелетные птицы в течение нескольких дней или ночей могли бы долететь в тропики. Однако птицы не могут сохранить указанную скорость пролета дольше, чем на несколько часов; они почти никогда не летят несколько дней или ночей подряд; как правило, их полет прерывается для короткого отдыха или для более длительных остановок; последние придают перелету в целом характер неторопливой "прогулки". При рассмотрении точно установленных кольцеванием средних скоростей дневного или ночного перелета отдельных видов всегда нужно иметь в виду, что они не характеризуют способность к полету и развиваемую на пролете скорость, а указывают лишь на длительность пролета и расстояние между местами кольцевания и находок окольцованных птиц в пересчете на один день. Многочисленные находки окольцованных птиц доказывают, что птицы быстро пролетают большую часть пути, а остальное время используют для отдыха в богатых кормом местах. Этот вид пролета встречается наиболее часто. Значительно реже наблюдается равномерное распределение нагрузки и отдыха. У птиц, летящих на большое расстояние, средний дневной путь равен примерно 150-200 км, в то время как летящие не так далеко не покрывают за это же время и 100 км. С этими данными согласуется продолжительность перелета в 2-3 или 3-4 мес. многих видов, которые зимуют в Тропической и Южной Африке. Так, например, аист, обычно отлетающий из Германии в конце августа, достигает зимовок в Южной Африке только в конце ноября или в декабре. При оценке дневного пути и скорости пролета нельзя упускать из виду и другой важный фактор: любые цифровые данные можно вычислить только для идеального пути перелета, т. е. для прямой линии, соединяющей места кольцевания и находки окольцованной птицы. В действительности пролетный путь всегда больше, отклонения от прямой часто весьма значительны и выполненная работа и скорость значительно выше вычисленных. Эти ошибки практически невозможно исключить, и поэтому их необходимо учитывать, особенно при очень длительных перелетах. Кроме того, следует обращать внимание и на то, когда были получены эти данные. Дело в том, что при весеннем перелете показатели во многих случаях значительно выше, чем при осеннем. В единичных случаях можно было бы с уверенностью доказать, что весенний перелет проходит вдвое быстрее осеннего, например у аиста, американского веретенника и жулана. Штреземан (1944) точно установил, что весной пролет жулана длится приблизительно 60 дней, а осенью - около 100 дней. В среднем эти птицы пролетают около 200 км в сутки. Однако они летят только ночью в течение 10 час. со скоростью 50 км/час. После такого пролета они всегда отдыхают, так что расстояние 1000 км покрывается ими за 5 суток: миграция - 2 ночи, сон - 3 ночи, кормежка - 5 дней. Еще несколько слов о максимальных скоростях и длительности пролета, характеризующих возможности перелетных птиц: камнешарка, небольшая прибрежная птица, окольцованная на Гельголанде, была найдена через 25 час. в Северной Франции, на 820 км южнее. Многочисленные мелкие певчие птицы регулярно перелетают за 12-15 час. Мексиканский залив шириной 750-1000 км. Согласно Моро (1938), некоторые небольшие сокола (Falco concolor и F. amurensis), а также азиатские щурки (Merops persicus и М. apiaster), зимующие на побережье Южной Африки, также пролетают не меньше 3000 км над морем. Гавайские острова служат местом, зимовки для ряда северных куликов, которые, мигрируя от Алеутских островов и Аляски, где расположены их гнездовья, вынуждены пролетать 3300 км над открытым. К пролету над водными пространствами можно приравнять пролет над большими пустынями. Такой пролег также, несомненно, проходит без перерыва, например пролет над Западной Сахарой мелких певчих птиц, трясогузок и коньков, требующий 30-40 час. непрерывной работы, если скорость их пролета считать равной приблизительно 50 км/час. |
 Обыкновенная пустельга может также двигаться по инерции, сложив крылья, или лететь при неполном их раскрытии.
Обыкновенная пустельга может также двигаться по инерции, сложив крылья, или лететь при неполном их раскрытии.
 Мало того, памятник почтовому голубю есть почти в каждом европейском городе. Большая их часть связана с заслугами почтовых голубей во время Второй мировой войны.
Мало того, памятник почтовому голубю есть почти в каждом европейском городе. Большая их часть связана с заслугами почтовых голубей во время Второй мировой войны. Этот факт был установлен благодаря датчикам слежения, прикрепленным к лапе одной из птиц этого вида. Эти данные даже зафиксированы в Книге рекордов Гиннеса.
Этот факт был установлен благодаря датчикам слежения, прикрепленным к лапе одной из птиц этого вида. Эти данные даже зафиксированы в Книге рекордов Гиннеса. Удар он наносит сложенными и прижатыми к туловищу лапами. По точным вычислениям, падая на добычу под углом в 25°, сокол летит со скоростью 75 метров в секунду; при падении под углом, близким к прямому, скорость увеличивается до 100 метров в секунду или 360 км/ч. Есть данные, что сапсан способен развить скорость до 440 км/ч, что сравнимо со скоростью некоторых самолётов. Сапсаны начинают преследование добычи уже с расстояния от одного до полутора километров.
Удар он наносит сложенными и прижатыми к туловищу лапами. По точным вычислениям, падая на добычу под углом в 25°, сокол летит со скоростью 75 метров в секунду; при падении под углом, близким к прямому, скорость увеличивается до 100 метров в секунду или 360 км/ч. Есть данные, что сапсан способен развить скорость до 440 км/ч, что сравнимо со скоростью некоторых самолётов. Сапсаны начинают преследование добычи уже с расстояния от одного до полутора километров. (Сравните: максимальная скорость экспресса - 39 м/сек, или 140 км/час.) Это, конечно, не означает, что птицы вообще не умеют летать быстрее. Гоняющиеся друг за другом стрижи развивают скорость до 200 км/час, а сокол бросается на жертву со скоростью 70 м/сек, т. е. 250 км/час. Но эти предельные скорости в течение очень короткого времени являются исключениями: они в лучшем случае характеризуют способность к полету некоторых видов, но их нельзя использовать для оценки скорости полета при миграциях, когда необходимо длительное напряжение.
(Сравните: максимальная скорость экспресса - 39 м/сек, или 140 км/час.) Это, конечно, не означает, что птицы вообще не умеют летать быстрее. Гоняющиеся друг за другом стрижи развивают скорость до 200 км/час, а сокол бросается на жертву со скоростью 70 м/сек, т. е. 250 км/час. Но эти предельные скорости в течение очень короткого времени являются исключениями: они в лучшем случае характеризуют способность к полету некоторых видов, но их нельзя использовать для оценки скорости полета при миграциях, когда необходимо длительное напряжение. Учитывая задерживающее или ускоряющее влияние ветра, можно точно измерить собственную скорость птиц на коротких расстояниях и в соответствии с этим вычислить истинную скорость пролета. Впервые такие расчеты произвел Тинеман на Курской косе. Впоследствии они были сделаны Мейнертцхагеном, Гаррисоном и др
Учитывая задерживающее или ускоряющее влияние ветра, можно точно измерить собственную скорость птиц на коротких расстояниях и в соответствии с этим вычислить истинную скорость пролета. Впервые такие расчеты произвел Тинеман на Курской косе. Впоследствии они были сделаны Мейнертцхагеном, Гаррисоном и др В общем она, очевидно, равна 40-80 км/час, причем скорость мелких певчих птиц приближается к низшим цифрам. Птицы, совершающие перелет ночью, летят, по-видимому, быстрее, чем мигрирующие днем. Бросается в глаза малая скорость миграции хищных и других крупных птиц. Одни и те же виды птиц летают в области гнездования обычно значительно медленнее, чем на пролете, если вообще эти скорости можно сравнивать.
В общем она, очевидно, равна 40-80 км/час, причем скорость мелких певчих птиц приближается к низшим цифрам. Птицы, совершающие перелет ночью, летят, по-видимому, быстрее, чем мигрирующие днем. Бросается в глаза малая скорость миграции хищных и других крупных птиц. Одни и те же виды птиц летают в области гнездования обычно значительно медленнее, чем на пролете, если вообще эти скорости можно сравнивать. Так возникают длительные миграции.
Так возникают длительные миграции. Эти же сроки относятся и к жулану. Ласточки мигрируют быстрее - с сентября до начала ноября. Сколь, однако, велики в данном случае индивидуальные различия, можно видеть на примере 3 окольцованных горихвосток-лысушек, одна из которых покрывала ежедневно 167 км, другая - 61 км и третья - лишь 44 км, причем эти числа уменьшаются по мере увеличения отрезка времени, для которого их вычисляют (6, 30 и 47 дней). Основываясь на этих результатах, можно сделать вывод, что дневная скорость более всего соответствует истинной скорости перелета в том случае, когда она вычисляется на основе общих показателей за короткий срок. Этот вывод лучше всего доказывают следующие примеры скорости пролетов отдельных птиц: аист покрыл за 2 дня 610 км, славка-черноголовка за 10 дней - 2200 км, лысуха за 7 дней - 1300 км, другая лысуха за 2 дня - 525 км, кряква за 5 дней - 1600 км. Этим данным можно противопоставить дневную скорость певчего дрозда - 40 км (вычислена за 56 дней перелета), зяблика - 17,4 км (вычислена за 23 дня перелета) и ястреба-перепелятника - 12,5 км (вычислена за 30 дней перелета).
Эти же сроки относятся и к жулану. Ласточки мигрируют быстрее - с сентября до начала ноября. Сколь, однако, велики в данном случае индивидуальные различия, можно видеть на примере 3 окольцованных горихвосток-лысушек, одна из которых покрывала ежедневно 167 км, другая - 61 км и третья - лишь 44 км, причем эти числа уменьшаются по мере увеличения отрезка времени, для которого их вычисляют (6, 30 и 47 дней). Основываясь на этих результатах, можно сделать вывод, что дневная скорость более всего соответствует истинной скорости перелета в том случае, когда она вычисляется на основе общих показателей за короткий срок. Этот вывод лучше всего доказывают следующие примеры скорости пролетов отдельных птиц: аист покрыл за 2 дня 610 км, славка-черноголовка за 10 дней - 2200 км, лысуха за 7 дней - 1300 км, другая лысуха за 2 дня - 525 км, кряква за 5 дней - 1600 км. Этим данным можно противопоставить дневную скорость певчего дрозда - 40 км (вычислена за 56 дней перелета), зяблика - 17,4 км (вычислена за 23 дня перелета) и ястреба-перепелятника - 12,5 км (вычислена за 30 дней перелета). Эти данные сравнимы с приведенными выше сведениями о горихвостках, на средние скорости которых при увеличении продолжительности пролета сильно влияют длительные остановки для отдыха.
Эти данные сравнимы с приведенными выше сведениями о горихвостках, на средние скорости которых при увеличении продолжительности пролета сильно влияют длительные остановки для отдыха.
 морем. Золотистой ржанке, особенно сильному летуну, для покрытия этого расстояния при скорости около 90 км/час потребовалось бы приблизительно 35 час. Более высокие скорости отмечены у другого вида ржанок, пролетающих от Новой Шотландии до северной оконечности Южной Америки 3600 км над морем. Почти невероятным кажется пролет одного из гнездящихся в Японии бекасов, который зимует в Восточной Австралии и должен покрыть почти 5000 км, чтобы достигнуть зимовок. В пути он, вероятно, совсем не отдыхает, так как в других местах его никогда не отмечали.
морем. Золотистой ржанке, особенно сильному летуну, для покрытия этого расстояния при скорости около 90 км/час потребовалось бы приблизительно 35 час. Более высокие скорости отмечены у другого вида ржанок, пролетающих от Новой Шотландии до северной оконечности Южной Америки 3600 км над морем. Почти невероятным кажется пролет одного из гнездящихся в Японии бекасов, который зимует в Восточной Австралии и должен покрыть почти 5000 км, чтобы достигнуть зимовок. В пути он, вероятно, совсем не отдыхает, так как в других местах его никогда не отмечали. Очень важно, чтобы ученики поняли, что
поддержание постоянной температуры тела
обеспечивается взаимодействием целого ряда
физиологических систем организма. Хорошее
знание этого материала необходимо для
объяснения сложных эволюционных и экологических
проблем.
Очень важно, чтобы ученики поняли, что
поддержание постоянной температуры тела
обеспечивается взаимодействием целого ряда
физиологических систем организма. Хорошее
знание этого материала необходимо для
объяснения сложных эволюционных и экологических
проблем. )
)
 Насекомоядные птицы наполняют свой желудок 5–6
раз в сутки, зерноядные – трижды.
Насекомоядные птицы наполняют свой желудок 5–6
раз в сутки, зерноядные – трижды. Для того
чтобы создать высокое артериальное давление,
сердце птиц должно сокращаться с большой силой и
высокой частотой (табл. 5).
Для того
чтобы создать высокое артериальное давление,
сердце птиц должно сокращаться с большой силой и
высокой частотой (табл. 5). Птицам пришлось развивать
различные приспособления и черты поведения,
позволяющие добывать достаточное количество
пропитания. Вот несколько примеров.
Птицам пришлось развивать
различные приспособления и черты поведения,
позволяющие добывать достаточное количество
пропитания. Вот несколько примеров.  Однако основным средством ориентации у ночных
сов служит не зрение, а слух. С его помощью сова по
писку и шороху узнает о присутствии грызунов и
точно определяет место нахождения добычи.
Однако основным средством ориентации у ночных
сов служит не зрение, а слух. С его помощью сова по
писку и шороху узнает о присутствии грызунов и
точно определяет место нахождения добычи.
 С помощью секундомера удалось определить и скорость полета сокола: 264-288 километров в час. Но вот насколько эти цифры точны - неизвестно. Многие
С помощью секундомера удалось определить и скорость полета сокола: 264-288 километров в час. Но вот насколько эти цифры точны - неизвестно. Многие Цапли и фазаны развивают скорость до 55-65, сойки - 40-48 километров в час. И что самое удивительное, неповоротливые индейки, оказывается, просто рекордсменки. Ведь они летают со скоростью 50-56 километров в час.
Цапли и фазаны развивают скорость до 55-65, сойки - 40-48 километров в час. И что самое удивительное, неповоротливые индейки, оказывается, просто рекордсменки. Ведь они летают со скоростью 50-56 километров в час.
 Теперь уже естественным ареалом ее обитания являются города, и намного реже это касается лесов.
Теперь уже естественным ареалом ее обитания являются города, и намного реже это касается лесов. Необходимо лишь несколько очень сильных взмахов крыльями и хотя бы небольшая возвышенность для того, чтобы легче было взлететь. Крылья сами по себе непропорционально большие, если все-таки их сравнивать с размерами самого тела.
Необходимо лишь несколько очень сильных взмахов крыльями и хотя бы небольшая возвышенность для того, чтобы легче было взлететь. Крылья сами по себе непропорционально большие, если все-таки их сравнивать с размерами самого тела. Но тем не менее, стриж не может быть лучше по маневренности полета, чем ласточка. Птица этого вида также отличается от стрижа строением лапок - у стрижа четыре пальца повернуты вперед, а у ласточек - три вперед и один назад. Именно поэтому они могут садиться на телеграфные провода и там легко удерживаться, а вот у стрижа подобное точно не получится.
Но тем не менее, стриж не может быть лучше по маневренности полета, чем ласточка. Птица этого вида также отличается от стрижа строением лапок - у стрижа четыре пальца повернуты вперед, а у ласточек - три вперед и один назад. Именно поэтому они могут садиться на телеграфные провода и там легко удерживаться, а вот у стрижа подобное точно не получится. Он хоть и не самая быстрая птица в мире, но состоит в Книге рекордов Гиннеса в связи со своими удивительными особенностями.
Он хоть и не самая быстрая птица в мире, но состоит в Книге рекордов Гиннеса в связи со своими удивительными особенностями. Скворцы могут развивать скорость приблизительно в 70 километров в час, и встречаются они на каждом континенте нашей планеты.
Скворцы могут развивать скорость приблизительно в 70 километров в час, и встречаются они на каждом континенте нашей планеты.Cкорости движения различных животных и птиц и расстояния, которые они проходят за время полета дроби до встречи. Все эти данные - итог исследований многих специалистов, и пренебрегать ими не следует. Конечно, не всякий сможет удержать в памяти столько цифр, вспомнить в нужный момент и правильно выбрать упреждение. Этого и не нужно. Главное - усвоить принцип определения правильного упреждения по различной дичи, идя на охоту, помнить характерные данные, потренироваться вхолостую, вскидывая ружье по мысленно представляемой цели. Таблица 42
Таблица 43
пожие материалы |

Особенности лётных качеств вороновых птиц Corvidae Текст научной статьи по специальности «Биологические науки»
ISSN 0869-4362
Русский орнитологический журнал 2008, Том 17, Экспресс-выпуск 420: 791-810
Особенности лётных качеств вороновых птиц Corvidae
Б. К. Штегман
К. Штегман
Второе издание. Первая публикация в 1954*
Вороновые птицы Corvidae представляют собой естественную группу, морфологически хорошо отграниченную от других семейств отряда воробьиных Passeriformes. Будучи самыми крупными представителями отряда, вороновые птицы существенным образом отличаются от прочих воробьиных характером полёта, в общем плавного, со сравнительно небыстрыми взмахами крыльев. Однако в пределах семейства лётные качества не однородны: уже на первый взгляд можно выделить две адаптивные группы, резко противопоставленные одна другой,— виды, населяющие открытый ландшафт, и лесные виды.
В качестве типичного представителя первой группы можно привести обыкновенную ворону (серую Corvus cornix и чёрную C. corone). Эту птицу все знают, но вряд ли широко известно, что она обладает большим летательным аппаратом. Действительно, размах крыльев у вороны около 1 м при среднем весе птицы около 0. 5 кг. Среднее отношение размаха к длине корпуса составляет 6.97, между тем как у гуся оно всего 4.9, у фазана — 3.7. Вместе с тем крылья широкие, так что площадь их очень значительная. В среднем ширина крыла составляет 143.7% длины корпуса. Вполне естественно, крыло вороны при большой ширине имеет тупую, округлую вершину. Это хорошо заметно на рисунке 1, так же как и пальцеобразное расхождение сильно развитых пропеллирующих маховых.
5 кг. Среднее отношение размаха к длине корпуса составляет 6.97, между тем как у гуся оно всего 4.9, у фазана — 3.7. Вместе с тем крылья широкие, так что площадь их очень значительная. В среднем ширина крыла составляет 143.7% длины корпуса. Вполне естественно, крыло вороны при большой ширине имеет тупую, округлую вершину. Это хорошо заметно на рисунке 1, так же как и пальцеобразное расхождение сильно развитых пропеллирующих маховых.
Последнее явление представляет собой закономерное следствие большой ширины и закруглённости крыла. В остроконечном крыле маховые, образующие вершину, быстро убывают в длине. При этом самую вершину образуют одно-два маховых, так что даже при совсем раскрытом крыле они полностью перекрываются. Если же крыло тупое, то вершину его образуют пять-шесть маховых, мало разнящихся по длине. При раскрытом крыле они расходятся веерообразно, в результате чего вершинные их части перестают взаимно перекрываться и разъединяются. Если такое крыло совершает взмахи, то каждое из разъединённых маховых в отдельности начинает пропеллировать. Сужение разъединённых вершин маховых является вторичным при-
Если такое крыло совершает взмахи, то каждое из разъединённых маховых в отдельности начинает пропеллировать. Сужение разъединённых вершин маховых является вторичным при-
* Штегман Б.К. 1954. Особенности лётных качеств вороновых птиц // Зоол. журн. 33, 3: 653-668.
способлением для улучшения их пропеллирующих качеств, в частности для придания им большей жёсткости, а также для сокращения переходной зоны между сплошной плоскостью крыла и разрезной его вершиной.
Таким образом, широкое тупое крыло вполне закономерно приводит к образованию пропеллирующих маховых, играющих существенную роль при активном полёте, в особенности при взлёте. У птиц с узкими крыльями пропеллирует вся вершинная часть крыла целиком, образуя во время опускания пронационный поворот в кисти. Но так как проксимальная часть крыла сохраняет положительный угол атаки, то в данный момент получается винтообразное перекручивание крыла, тем более сильное, чем больший пронационный поворот делает кисть.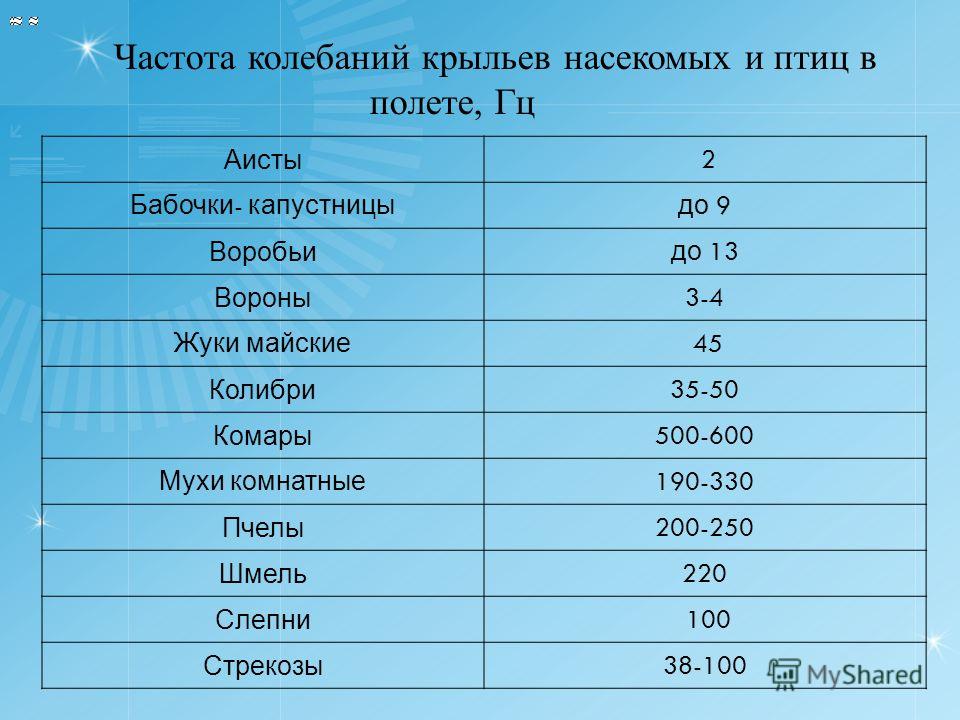 С увеличением ширины крыла способность его к перекручиванию (без разъединения маховых) уменьшается и пропеллирова-ние должно производиться всё в большей степени за счёт первых маховых. В результате мы видим удивительное взаимопроникновение различных принципов: в широком тупом крыле вершинные маховые по необходимости расходятся, но они по необходимости же используются как пропеллирующие элементы, так как широкое крыло не способно на сильное перекручивание и, следовательно, вершинная часть его целиком не может дать такой силы тяги, как это доступно для узкого крыла.
С увеличением ширины крыла способность его к перекручиванию (без разъединения маховых) уменьшается и пропеллирова-ние должно производиться всё в большей степени за счёт первых маховых. В результате мы видим удивительное взаимопроникновение различных принципов: в широком тупом крыле вершинные маховые по необходимости расходятся, но они по необходимости же используются как пропеллирующие элементы, так как широкое крыло не способно на сильное перекручивание и, следовательно, вершинная часть его целиком не может дать такой силы тяги, как это доступно для узкого крыла.
Из вышесказанного явствует, что для вороны, обладающей широкими крыльями, пропеллирующие маховые имеют большое функциональное значение. Действительно, они у вороны сильно развиты, и пропеллирующая, разрезная часть крыла у этой птицы занимает около одной трети длины его. Сужения на внешних опахалах заметны до 6-го махового, так что пропеллирующих маховых всего пять. Место расхождения маховых от 2-го до 6-го проходит по прямой, параллельной продольной оси тела. Эта особенность пропеллирующих маховых, встречающаяся и у других птиц и отмеченная в литературе, видимо, связана с необходимостью сокращения переходной зоны между сплошной и разрезной частями крыла. При подобном устройстве крыла про-пеллирующие части маховых начинаются на одинаковом расстоянии от тела и переходная зона имеет всюду одинаковую скорость движения при взмахах крыла.
Эта особенность пропеллирующих маховых, встречающаяся и у других птиц и отмеченная в литературе, видимо, связана с необходимостью сокращения переходной зоны между сплошной и разрезной частями крыла. При подобном устройстве крыла про-пеллирующие части маховых начинаются на одинаковом расстоянии от тела и переходная зона имеет всюду одинаковую скорость движения при взмахах крыла.
Впрочем, первое маховое у вороны, далеко не доходящее до вершины крыла, является пропеллирующим на всей своей видимой части, т.е. от вершин кроющих крыла, и к нему явно неприложим принцип, вероятный для устройства других пропеллирующих маховых. Какова его функция во время активного полёта — определить трудно, но положение его во время планирования наблюдать достаточно просто. Оказывается, оно в таких случаях бывает вынесено вперед и, бла-
годаря лётной нагрузке, загнуто вверх, выше переднего края самого крыла.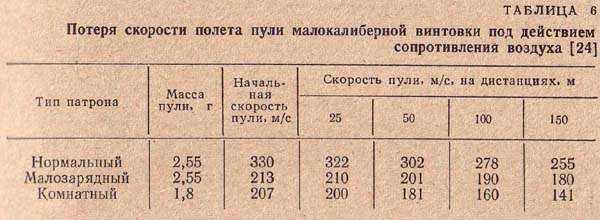 В этом положении его функция, несомненно, аналогична функции крылышка, аэродинамическое значение которого было не столь давно выяснено. Соответствуя предкрылку самолётов, крылышко у птиц даёт возможность увеличивать угол атаки и подъёмную силу крыла. Однако будучи сравнительно малым и коротким, оно простирает свое действие только на часть длины крыла. Первое маховое расположено дистальнее крылышка, и, таким образом, его действие является дополнением к действию последнего. Такое функциональное значение первого махового свойственно большинству вороновых птиц, а также, вероятно, и многим другим птицам, имеющим широкие и закруглённые крылья. В частности, у ястребов его значение, несомненно, велико. У этих птиц первое маховое даже в сложенном крыле имеет тенденцию ложиться поверх других маховых, а в распростёртом крыле ясно видно, что его действие начинается как раз от вершины крылышка.
В этом положении его функция, несомненно, аналогична функции крылышка, аэродинамическое значение которого было не столь давно выяснено. Соответствуя предкрылку самолётов, крылышко у птиц даёт возможность увеличивать угол атаки и подъёмную силу крыла. Однако будучи сравнительно малым и коротким, оно простирает свое действие только на часть длины крыла. Первое маховое расположено дистальнее крылышка, и, таким образом, его действие является дополнением к действию последнего. Такое функциональное значение первого махового свойственно большинству вороновых птиц, а также, вероятно, и многим другим птицам, имеющим широкие и закруглённые крылья. В частности, у ястребов его значение, несомненно, велико. У этих птиц первое маховое даже в сложенном крыле имеет тенденцию ложиться поверх других маховых, а в распростёртом крыле ясно видно, что его действие начинается как раз от вершины крылышка.
Итак, ворона имеет большие широкие крылья с хорошо развитыми пропеллирующими маховыми. Хвост у неё тоже достаточно большой, будучи в среднем на одну треть длиннее корпуса. Таким образом, у вороны несущие поверхности велики, удельная нагрузка на них мала, в связи с чем маховые и рулевые относительно мягкие.
Хвост у неё тоже достаточно большой, будучи в среднем на одну треть длиннее корпуса. Таким образом, у вороны несущие поверхности велики, удельная нагрузка на них мала, в связи с чем маховые и рулевые относительно мягкие.
Исходя из этих особенностей летательного аппарата вороны, можно предположить, что эта птица летает легко и экономно, способна на дальние перелёты, но не приспособлена к скоростному полету. Наблюдения вполне подтверждают это предположение. Действительно, ворона летит легко, с неторопливыми взмахами крыльев. Во время горизонтального полёта она делает в среднем 3 взмаха в секунду. Взлетает она с горизонтальной поверхности тоже легко, может подниматься достаточно круто, но делает это не быстро. Она без труда пролетает значительные расстояния, иногда многие десятки километров в день. Дальние перелёты ворона совершает преимущественно на большой высоте; снижается постепенно, делая более редкие взмахи и планируя на значительные расстояния. огаНв + т. supracoracoideum + т. delto-ideus) обеих сторон в процентах от веса тела для вороны в сравнении с
огаНв + т. supracoracoideum + т. delto-ideus) обеих сторон в процентах от веса тела для вороны в сравнении с
некоторыми другими птицами: ворона Corvus cornix - 16.54, голубь Columba livia - 26.14, вяхирь Columba palumbus - 28.9, рябок Pterocles alchata - 28.45, тиркушка Glareola pratincola - 21.66, кряква Anas platyrhynchos - 25, шилохвость Anas acuta - 23.7, фазан Phasianus col-chicus - 24.6.
Прочие наши вороновые птицы на первый взгляд кажутся очень сходными с обыкновенной вороной и по форме летательного аппарата, и по типу полёта. Однако при более точном сравнении различных видов оказывается, что вороний «стандарт» касается лишь общих черт, которые сходны для всех представителей группы, в частностях же наблюдается значительное разнообразие.
Одним из близких родственников вороны является грач Corvus frugilegus. Сравнение его с вороной особенно интересно потому, что эти виды примерно одинаковых размеров. Можно лишь сказать, что грач в общем несколько легче. По нашим измерениям, вес различных подвидов вороны (серой и чёрной) колеблется от 405 до 640 г, грача - от 400 до 544 г. Общее сходство во внешности между этими видами таково, что неоднократно начинающие орнитологи (а иногда и более опытные) определяли молодых грачей, у которых ещё имеются перья у основания клюва, как чёрных ворон Corvus corone. Конечно, в образе жизни и в поведении различия между этими видами достаточно велики, так что на воле их трудно спутать. В частности, опытный наблюдатель легко отличит грача от вороны и по картине полёта, и по большей скорости его.
Можно лишь сказать, что грач в общем несколько легче. По нашим измерениям, вес различных подвидов вороны (серой и чёрной) колеблется от 405 до 640 г, грача - от 400 до 544 г. Общее сходство во внешности между этими видами таково, что неоднократно начинающие орнитологи (а иногда и более опытные) определяли молодых грачей, у которых ещё имеются перья у основания клюва, как чёрных ворон Corvus corone. Конечно, в образе жизни и в поведении различия между этими видами достаточно велики, так что на воле их трудно спутать. В частности, опытный наблюдатель легко отличит грача от вороны и по картине полёта, и по большей скорости его.
Каковы же морфологические различия между этими видами? Оказывается, у грача крылья несколько длиннее, чем у вороны: отношение размаха к длине корпуса в среднем составляет 7.08. Вместе с тем ширина крыла у грача заметно меньше: 133.18% от длины корпуса против 143.73% у вороны. Эти особенности крыла грача хорошо заметны и при визуальном сравнении (рис. 2). В связи с меньшей шириной крыльев у грача и число пропеллирующих перьев меньше, чем у вороны. В то время как у последнего вида сужение внешнего опахала имеется до 6-го махового включительно, у грача оно заметно лишь до 5-го. Таким образом, число пропеллирующих маховых у вороны равно пяти, у грача - четырём. Наконец, по рисункам хорошо видно, что у грача хвост короче и значительно более закруглённый, чем у вороны. В цифрах разница в длине хвоста выражается тоже достаточно ясно: у вороны хвост составляет 132.1% от длины корпуса, у грача - 128.1%.
2). В связи с меньшей шириной крыльев у грача и число пропеллирующих перьев меньше, чем у вороны. В то время как у последнего вида сужение внешнего опахала имеется до 6-го махового включительно, у грача оно заметно лишь до 5-го. Таким образом, число пропеллирующих маховых у вороны равно пяти, у грача - четырём. Наконец, по рисункам хорошо видно, что у грача хвост короче и значительно более закруглённый, чем у вороны. В цифрах разница в длине хвоста выражается тоже достаточно ясно: у вороны хвост составляет 132.1% от длины корпуса, у грача - 128.1%.
Итак, мы видим, что у грача, по сравнению с вороной, летательный аппарат обнаруживает приспособление к более скоростному полёту. Меньшее количество пропеллирующих маховых у грача, несомненно, компенсируется тем, что более узкое его крыло способно к более сильному винтообразному перекосу, следовательно, его вершинная часть
целиком в состоянии более сильно пропеллировать. Более короткий и сильно закруглённый хвост способствует большей аэродинамичности летящего грача, так же как и меньшая ширина крыльев. Однако и то и другое приводит к уменьшению несущей поверхности грача, в результате чего его полёт, будучи более быстрым, чем у вороны, должен быть и менее экономным. Действительно, летательная мускулатура у грача, более мощная, чем у вороны, составляет в среднем 18.52% от веса тела. Конечно, интенсивная работа более мощной летательной мускулатуры должна предъявлять и большие требования к деятельности сердца. И действительно, вес сердца у грача составляет в среднем 11.96% от веса тела против 10.6% у вороны.
Более короткий и сильно закруглённый хвост способствует большей аэродинамичности летящего грача, так же как и меньшая ширина крыльев. Однако и то и другое приводит к уменьшению несущей поверхности грача, в результате чего его полёт, будучи более быстрым, чем у вороны, должен быть и менее экономным. Действительно, летательная мускулатура у грача, более мощная, чем у вороны, составляет в среднем 18.52% от веса тела. Конечно, интенсивная работа более мощной летательной мускулатуры должна предъявлять и большие требования к деятельности сердца. И действительно, вес сердца у грача составляет в среднем 11.96% от веса тела против 10.6% у вороны.
Рис. 1. Серая ворона Согуив согпіх.
Рис. 2. Грач Согуив іїидіівдив.
Из вышеприведённого сравнения явствует, что такие на первый взгляд близкие виды, как ворона и грач, обладают значительными различиями в приспособлениях к полёту. Ворона приспособлена к более медленному и экономному полёту, грач — к более быстрому полёту, требующему, однако, большей затраты энергии. Сильное развитие мускулатуры и большой вес сердца у грача указывают на то, что эта птица хорошо приспособлена к более быстрому полёту и может совер-
Ворона приспособлена к более медленному и экономному полёту, грач — к более быстрому полёту, требующему, однако, большей затраты энергии. Сильное развитие мускулатуры и большой вес сердца у грача указывают на то, что эта птица хорошо приспособлена к более быстрому полёту и может совер-
шать значительные перелёты. Впрочем, уже само приспособление к скоростному полёту включает в себя и приспособление к дальним перелётам. Если исключить хищников, способных на кратковременное ускорение полёта (чаще путем пикирования) для того, чтобы настигнуть добычу, то скоростной полёт может иметь явные преимущества лишь при передвижении на дальние дистанции. При небольших перелётах разница в скорости практически мало сказывается.
Установив особенности лётных качеств грача, конечно, желательно определить и биологическое значение их. И тут невольно напрашивается следующее соображение. Грач на большей части своего ареала является перелётной птицей, причём иногда отлетает далеко. Ворона на значительной части своего ареала оседла или производит сезонные кочёвки. Лишь местами она совершает настоящие перелёты, но и то на меньшие расстояния, чем грач. Правда, за последнее время неоднократно высказывались сомнения в том, что перелёты предъявляют повышенные требования к лётным способностям птиц. При этом в качестве основного довода указывалось на то, что большинство птиц в гнездовой период «у себя дома» пролетает в течение дня в общей сложности не меньшее число километров, чем во время перелёта. Разница заключается лишь в том, что в первом случае совершаются перелёты в разных направлениях с возвратом неизменно к одной точке, во втором же случае птица всё свое дневное расстояние пролетает в одном направлении.
Ворона на значительной части своего ареала оседла или производит сезонные кочёвки. Лишь местами она совершает настоящие перелёты, но и то на меньшие расстояния, чем грач. Правда, за последнее время неоднократно высказывались сомнения в том, что перелёты предъявляют повышенные требования к лётным способностям птиц. При этом в качестве основного довода указывалось на то, что большинство птиц в гнездовой период «у себя дома» пролетает в течение дня в общей сложности не меньшее число километров, чем во время перелёта. Разница заключается лишь в том, что в первом случае совершаются перелёты в разных направлениях с возвратом неизменно к одной точке, во втором же случае птица всё свое дневное расстояние пролетает в одном направлении.
Однако эти соображения могут быть признаны правильными лишь для кочующих птиц, как, например, синицы, или для таких воздухо-реев, как стрижи и ласточки. Для большинства же перелётных птиц перелёт, безусловно, предъявляет повышенные требования к лётным способностям. Не говоря уже о том, что во многих случаях дневные «порции» полёта повышены по сравнению с перелётами в гнездовый период, главная разница заключается в том, что перелёт требует передвижения на дальние дистанции без передышки.
Не говоря уже о том, что во многих случаях дневные «порции» полёта повышены по сравнению с перелётами в гнездовый период, главная разница заключается в том, что перелёт требует передвижения на дальние дистанции без передышки.
Таким образом, мы можем без особых погрешностей морфологические особенности грача и вытекающие из них особенности его лётных качеств привести в связь с его сезонными миграциями. Но обладая способностью к дальним перелётам, грачи пользуются ими и в гнездо-вый период. В частности, эти птицы при случае гнездятся далеко от своих кормовых угодий. Такие гнездовья приходилось видеть, например, в центре Ленинграда, откуда грачам приходится летать за кормом минимум за 8-10 км. В период выкармливания птенцов в таких колониях родители ежедневно пролетают по нескольку сотен километров, что, в особенности в ветреную погоду, должно представлять громадную затрату энергии.
Более сильно отличается от вороны галка monedula, выде-
ляемая большинством орнитологов в особый род.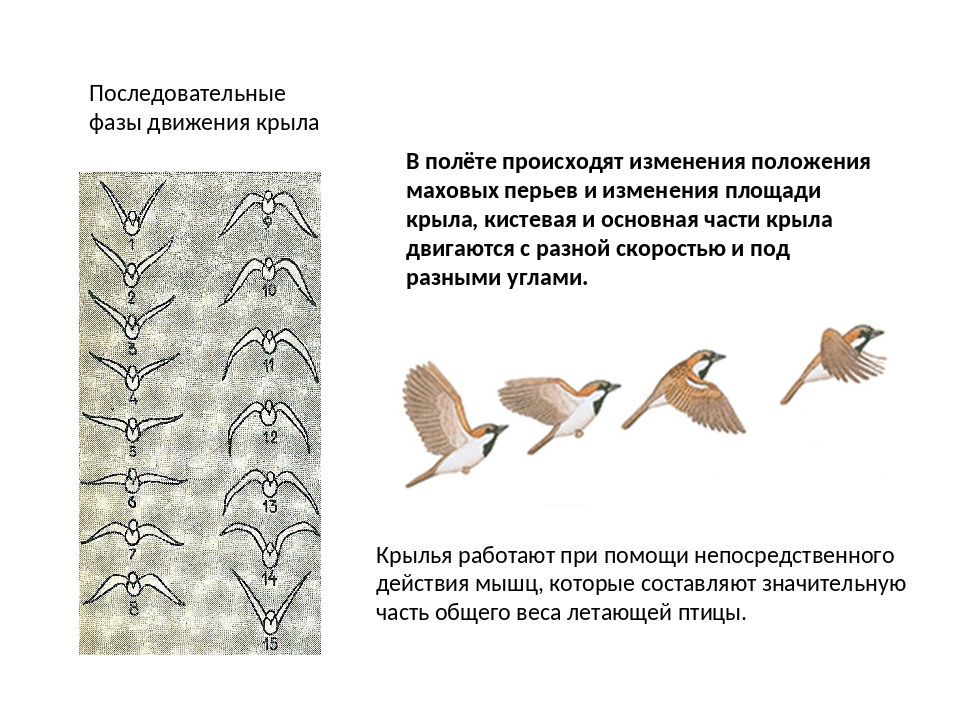 Вес этой птицы в среднем около 200 г, т.е. в два с лишним раза меньше веса вороны, что необходимо учитывать при сравнении летательного аппарата обоих видов. Известно, что при изменении размеров тела поверхность и с ней лобовое сопротивление изменяются в квадрате к линейным измерениям, а масса — в кубе. Следовательно, с уменьшением размеров птицы её относительное лобовое сопротивление возрастает, так что для сохранения одинаковой скорости полёта маленьким птицам более важно иметь особо обтекаемые формы, чем более крупным.
Вес этой птицы в среднем около 200 г, т.е. в два с лишним раза меньше веса вороны, что необходимо учитывать при сравнении летательного аппарата обоих видов. Известно, что при изменении размеров тела поверхность и с ней лобовое сопротивление изменяются в квадрате к линейным измерениям, а масса — в кубе. Следовательно, с уменьшением размеров птицы её относительное лобовое сопротивление возрастает, так что для сохранения одинаковой скорости полёта маленьким птицам более важно иметь особо обтекаемые формы, чем более крупным.
Рис. 3. Галка Со!оеив топеби!а.
Рис. 4. Ворон Согуив согах.
Уже при первом взгляде на рисунок галки (рис. 3) бросается в глаза, что форма крыльев у неё существенно иная, чем у вороны: картина полёта галки производит явно более стройное впечатление, что создаётся в основном более узкими и менее закруглёнными крыльями. Действительно, у галки ширина крыла составляет в среднем 125.83% длины тела, что значительно меньше, чем у вороны. Число пропелли-рующих маховых такое же, как и у грача, но сужения маховых начинаются значительно ближе к их вершинам, так что пропеллирующие части более короткие и разрезная часть крыла мало бросается в глаза.
Действительно, у галки ширина крыла составляет в среднем 125.83% длины тела, что значительно меньше, чем у вороны. Число пропелли-рующих маховых такое же, как и у грача, но сужения маховых начинаются значительно ближе к их вершинам, так что пропеллирующие части более короткие и разрезная часть крыла мало бросается в глаза.
Кроме того, крыло более острое. У вороны и грача 3-е и 4-е маховые почти одинаковой длины, у галки 4-е заметно короче 3-го а последующие быстро убывают в длине.
Наконец, у галки крылья заметно короче, чем у вороны. На рисунке эта разница не бросается в глаза, так как ширина крыльев невелика и хвост тоже короче, чем у вороны. На самом деле отношение размаха к длине корпуса составляет всего 6.53, т.е. на полкорпуса меньше, чем у вороны. Длина хвоста составляет в среднем 118.7% длины корпуса, что тоже значительно меньше, чем у вороны.
Таким образом, у галки несущие поверхности значительно меньше, чем у двух предыдущих видов. Крылья короче, уже, более заострены, хвост короче, и эти особенности летательного аппарата, улучшающие его аэродинамические качества, столь значительны, что, несомненно, не только компенсируют малые размеры галки, но должны бы ещё и увеличить скорость её полёта. Действительно, галка летает быстро, очень заметно превосходя в этом отношении ворону и грача. Взмахи крыльев у неё довольно быстрые, около четырёх в секунду, но ясно видно, что галка от полёта не устаёт, с лёгкостью перелетает на большие расстояния, часто подолгу играет в воздухе, без явной надобности поднимаясь на большую высоту. От гнездовий она часто очень далеко летает за кормом. В значительной части своего ареала галка совершает сезонные перелёты.
Крылья короче, уже, более заострены, хвост короче, и эти особенности летательного аппарата, улучшающие его аэродинамические качества, столь значительны, что, несомненно, не только компенсируют малые размеры галки, но должны бы ещё и увеличить скорость её полёта. Действительно, галка летает быстро, очень заметно превосходя в этом отношении ворону и грача. Взмахи крыльев у неё довольно быстрые, около четырёх в секунду, но ясно видно, что галка от полёта не устаёт, с лёгкостью перелетает на большие расстояния, часто подолгу играет в воздухе, без явной надобности поднимаясь на большую высоту. От гнездовий она часто очень далеко летает за кормом. В значительной части своего ареала галка совершает сезонные перелёты.
В связи с большой скоростью полёт у галки должен быть относительно мало экономным, вследствие чего можно предположить, что летательная мускулатура у неё мощная. Действительно, вес летательной мускулатуры в среднем составляет 18. 61% веса тела, что даже ещё немного больше, чем у грача. Что же касается сердца, то оно значительно крупнее, чем у предыдущих видов, составляя в среднем 13.27% веса тела. Эта последняя особенность галки, вероятно, связана с особой живостью и подвижностью данной птицы, о чём уже упоминалось.
61% веса тела, что даже ещё немного больше, чем у грача. Что же касается сердца, то оно значительно крупнее, чем у предыдущих видов, составляя в среднем 13.27% веса тела. Эта последняя особенность галки, вероятно, связана с особой живостью и подвижностью данной птицы, о чём уже упоминалось.
Впрочем, нет основания полагать, что галка является видом, узко приспособленным к скоростному полёту. Выше перечисленные качества её вполне относительны. Достаточно сравнить галку хотя бы с сизым голубем, живущим рядом с ней в поселениях и мало отличающимся от неё по величине, чтобы в этом убедиться. У голубя, более узко приспособленного к скоростному полёту, размах крыльев всего в 6 раз больше длины корпуса, крылья значительно более узкие, с острой компактной вершиной. По сравнению с голубем, у галки крылья большие, хвост длинный, а полёт менее стремительный.
Иным комплексом признаков отличается ворон Corvus corax — самый крупный представитель семейства, вес которого составляет примерно 1 кг.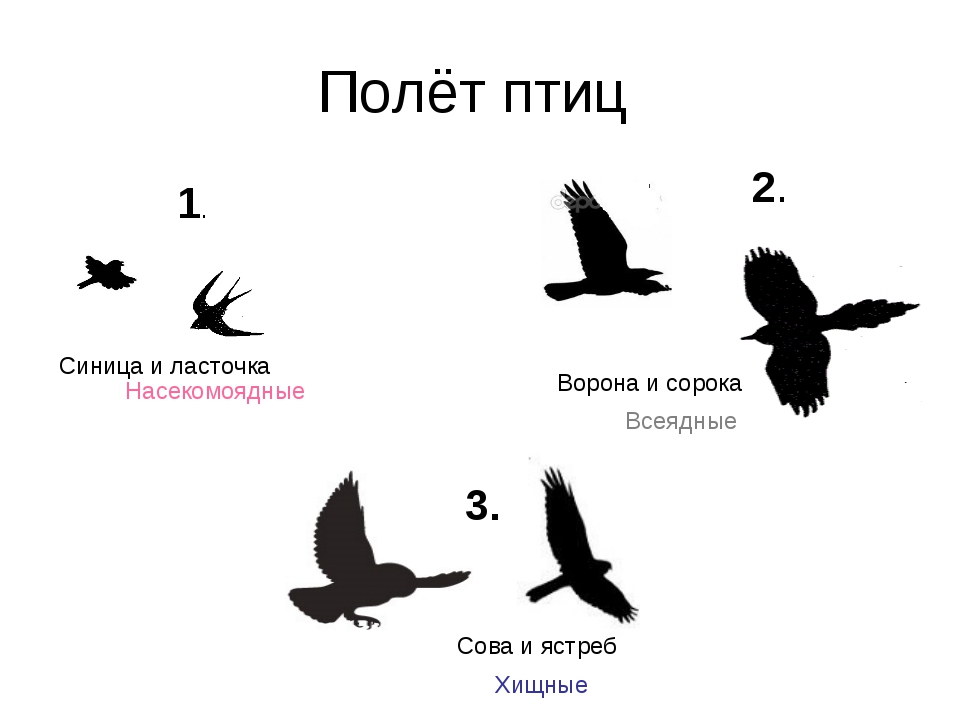 Исходя из того, что ворон по крайней мере вдвое тяжелее вороны, можно было бы предположить у него достаточно широкие
Исходя из того, что ворон по крайней мере вдвое тяжелее вороны, можно было бы предположить у него достаточно широкие
крылья, но на самом деле это не так. Ширина крыла у ворона составляет всего 123.41% длины корпуса, примерно как у галки. Между тем размах крыльев в 7.36 раза длиннее корпуса, что значительно больше, чем у всех ранее рассмотренных видов. Таким образом, у ворона крылья длинные и узкие. Вершина крыла закруглённая, и пропелли-рующие маховые сильно развиты, но число их такое же, как у грача и галки, так как сужения внешних опахал имеются только до 5-го махового (рис. 4). Вершины пропеллирующих маховых заострены, чем ворон отличается от прочих видов семейства, и, кроме того, относительно жёсткие. Длина хвоста у ворона несколько больше, чем у вороны, составляя 134.6% длины корпуса, форма его резко клиновидная, чего у других — настоящих — ворон не встречается.
Из вышеприведённого описания явствует, что ворон, по сравнению с вороной, приспособлен к более быстрому полёту. .
.
Наблюдения над образом жизни ворона вполне подтверждают предположения об особенностях его полёта, сделанных на основе изучения его морфологии. Действительно, ворон летает значительно быстрее вороны, делает часто дальние перелёты, поднимаясь при этом на громадную высоту. Вместе с тем он, несмотря на крупные размеры, отличается большой ловкостью полёта, в частности хорошо пикирует, напоминая в этом отношений хищных птиц. Их же он напоминает и способностью к парению, которой часто пользуется, в особенности в горных местах и в пустынях, в которых за день поверхность земли сильно прогревается. Найдя восходящее воздушное течение, он при случае подолгу носится без единого взмаха крыльев, часто при этом ещё набирая высоту. Следует отметить, что ранее перечисленные виды вороновых не парят. Только один раз мне пришлось видеть пару грачей, паривших в жаркий солнечный день над песками южного Прибалхашья. Для ворона же парение — обычный тип полёта.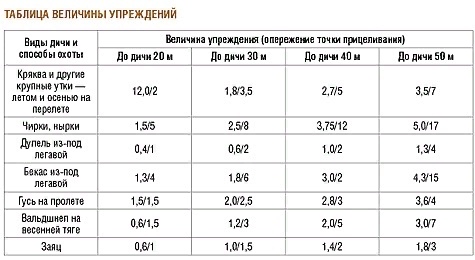
Указанные особенности полёта ворона недаром напоминают хищных птиц. По своему образу жизни, в частности питанию, ворон в большей степени «хищный», чем все его родственники, в связи с чем и
выработались характерные качества его полёта. Быстрота полёта и способность к дальним перелётам, видимо, связаны с крупными размерами ворона и большим количеством потребляемой им пищи.
Вес летательной мускулатуры у ворона примерно такой же, как у вороны, но сердце более тяжёлое, составляя 12.1% общего веса. Впрочем, измерен был лишь 1 экз., чего для каких-либо выводов, конечно, недостаточно.
Таковы в общих чертах особенности лётных качеств и морфологических особенностей настоящих ворон, населяющих в основном открытый ландшафт. Зачастую полёт этих видов расценивается как неплохой. Вторая группа вороновых птиц с точки зрения качества полёта расценивается менее положительно. В качестве одного из представителей этих «плохо» летающих вороновых птиц можно рассмотреть обыкновенную сойку Оагги1ш glandarius.
В качестве одного из представителей этих «плохо» летающих вороновых птиц можно рассмотреть обыкновенную сойку Оагги1ш glandarius.
Сойка - настоящая лесная птица, редко показывающаяся в открытых местах, От ранее описанных вороновых птиц она резко отличается по форме летательного аппарата. Длина крыльев значительно меньше, чем у всех предыдущих видов, и размах превышает длину корпуса всего в 5.65 раза. Зато ширина крыльев очень велика, составляя 154.3% длины корпуса. Пропеллирующие маховые сильно развиты, и число их равно 5, так как ещё 6-е маховое имеет сужение внешнего опахала. Вершина крыла более сильно закруглена, чем у настоящих ворон, так как 3-е, 4-е и 5-е маховые почти одинаковой длины. Вообще, удлинение крыла ничтожно, так что в распростёртом виде форма приближается к округлой. Хвост длинный и прямо срезанный, длина его составляет 157.5% длины корпуса.
В общем сойка во время полёта производит очень своеобразное впечатление, и кажется, что крылья у неё необыкновенно короткие (рис. 5). На самом деле это не так. Если сравнить сойку с близкой к ней по величине галкой, то, конечно, бросается в глаза громадная разница в форме крыльев. Однако если у сойки крылья короче, то ширина их значительно больше, так что площадь их не меньше, чем у галки. Хвост значительно длиннее, чем у галки.
5). На самом деле это не так. Если сравнить сойку с близкой к ней по величине галкой, то, конечно, бросается в глаза громадная разница в форме крыльев. Однако если у сойки крылья короче, то ширина их значительно больше, так что площадь их не меньше, чем у галки. Хвост значительно длиннее, чем у галки.
Еще более поучительно сравнение сойки с голубями, приспособленными к скоростному полёту. Крылья голубей производят впечатление более длинных. Между тем у горлицы, близкой по величине к сойке, отношение размаха к длине корпуса 5.6, т.е. примерно такое же, как у сойки, ширина же крыла значительно меньше, так что и площадь крыла очень заметно меньше, чем у сойки. Таким образом, понятие «длинное крыло» очень относительно и зависит в значительной мере от степени удлинения его, т.е. от отношения длины крыла к его ширине.
Всё-таки, по сравнению с ранее описанными видами, у сойки крылья укорочены, что, несомненно, является приспособлением к полёту среди древесной чащи. Чем меньше размах крыльев, тем легче делать резкие повороты. Чем меньше поступательная скорость, тем меньше центробежная сила, развивающаяся при повороте. Следовательно, для
Чем меньше размах крыльев, тем легче делать резкие повороты. Чем меньше поступательная скорость, тем меньше центробежная сила, развивающаяся при повороте. Следовательно, для
Рис. б. Сойка Garrulus glandarius.
Рис. б. Сорока Pica pica.
полёта в лесу необходимо иметь небольшой размах, но большую площадь крыльев, создающую большую подъёмную силу и позволяющую держаться в воздухе на небольших скоростях. Это может быть достигнуто лишь увеличением ширины крыльев. Большая длина хвоста тоже содействует маневренности полёта и поэтому полезна для птиц, летающих среди древесных зарослей.
Приспособленная к полёту среди лесной чащи, сойка передвигается не быстро и почти бесшумно, она пролетает через самые густые заросли, то планируя, то несколькими быстрыми взмахами давая себе
новый импульс, делая резкие повороты в любом направлении, внезапно тормозя полёт и так же быстро приобретая опять нужную скорость. Большую роль в этом способе передвижения играет способность сойки в любой момент схватиться за ветку и сесть; ловко перескакивая с ветки на ветку, выбраться из чащи, где нельзя расправить крыльев, и, оттолкнувшись, сразу полететь дальше. Такой способ передвижения для ранее описанных настоящих ворон совсем недоступен.
Большую роль в этом способе передвижения играет способность сойки в любой момент схватиться за ветку и сесть; ловко перескакивая с ветки на ветку, выбраться из чащи, где нельзя расправить крыльев, и, оттолкнувшись, сразу полететь дальше. Такой способ передвижения для ранее описанных настоящих ворон совсем недоступен.
Но эти качества полёта сойки развились в ущерб другим качествам его. Большая ширина крыльев сойки создает большое сопротивление, вследствие чего скорость полёта этой птицы очень невелика. Следовательно, бороться с сильными ветрами сойка не может, тем более, что широкое крыло — относительно мягкое и задний край его при ускорении движения легко начинает вибрировать. Перелетая через открытые пространства, сойка движется странным, неровным полётом, в котором быстрые и медленные взмахи чередуются. На более дальние расстояния сойка передвигается цепью траекторий, которая создаётся тем, что после каждого взмаха птица проносится какое-то расстояние с прижатыми к телу крыльями. Этот тип полёта напоминает полёт мелких птиц, которые тоже передвигаются траекториями. Видимо, как и у мелких птичек, у сойки крылья создают столь сильное сопротивление воздуха, что для неё выгоднее какое-то расстояние пролетать с прижатыми крыльями, нежели планировать. Но такой полёт требует очень быстрых и сильных взмахов, и поэтому сойка неохотно перелетает на дальние расстояния. Перелёт, который галка совершает играя, для сойки может оказаться не только затруднительным, но даже и невыполнимым без отдыха.
Этот тип полёта напоминает полёт мелких птиц, которые тоже передвигаются траекториями. Видимо, как и у мелких птичек, у сойки крылья создают столь сильное сопротивление воздуха, что для неё выгоднее какое-то расстояние пролетать с прижатыми крыльями, нежели планировать. Но такой полёт требует очень быстрых и сильных взмахов, и поэтому сойка неохотно перелетает на дальние расстояния. Перелёт, который галка совершает играя, для сойки может оказаться не только затруднительным, но даже и невыполнимым без отдыха.
Качества полёта других лесных ворон - кукши Perisoreus infaustus и голубой сороки Cyanopica cyanus очень сходны с таковыми сойки. Близка к этим птицам по лётным способностям и обыкновенная сорока Pica pica. Эта птица тоже приспособлена к полёту в лесу и плохо летает на дальние расстояния. Совершая более далекий перелёт, сорока тоже странно дёргается в воздухе, работая крыльями то медленнее, то быстрее, и скоро отстаёт от рядом летящей вороны, хотя явно затрачивает больше энергии, чем последняя. Среди древесной чащи она ловко лавирует, причём, несомненно, большую пользу ей приносит её ступенчатый хвост, длина которого более чем в 2 раза превышает длину корпуса.
Среди древесной чащи она ловко лавирует, причём, несомненно, большую пользу ей приносит её ступенчатый хвост, длина которого более чем в 2 раза превышает длину корпуса.
Но, по сравнению с сойкой, сорока, безусловно, чаще перелетает через открытые пространства и вообще способна на более дальние перелёты. В то время как сойка является настоящей лесной птицей, редко спускающейся на землю, сорока населяет преимущественно опушки, перелески и особенно характерна для лесостепи. Значительную часть своей пищи она собирает на земле, по которой хорошо ходит, как и на-
стоящие вороны, к которым она, несомненно, очень близка. В соответствии с особенностями образа жизни и летательный аппарат сороки приобрёл некоторые специфические особенности. Размах крыльев несколько больше, ширина крыльев заметно меньше, в особенности вершинная часть более узкая (рис. 6). Кроме того, у сороки крыло менее закруглённое, чем у сойки. У последней 3-е, 4-е, 5-е и 6-е маховые почти одинаковой длины, у первой 5-е и, тем более, 6-е маховые значительно короче 3-го и 4-го. Таким образом, получается впечатление, что крылья сороки менее узко приспособлены к полёту среди древесных зарослей, чем у сойки, но зато в какой-то степени допускают более быстрый полёт, а также перелёты на более дальние расстояния. Впрочем, у сороки внутривидовая изменчивость значительна; в частности, и летательный аппарат изменяется достаточно сильно. Так, например, у североафриканской формы P. p. mauretanica (Malherbe, 1845) крылья заметно короче, а хвост длиннее, чем у наших подвидов, что заставляет предполагать большую привязанность этой формы к лесу. С другой стороны, центральноазиатский подвид P. p. bottanensis (Delessert, 1840) имеет значительно более длинные крылья и более короткий хвост. Он населяет высокогорье, в частности субальпийскую зону. В этих местах сорока, перелетая через глубокие долины, совершает то и дело сравнительно дальние и высокие полёты.
У последней 3-е, 4-е, 5-е и 6-е маховые почти одинаковой длины, у первой 5-е и, тем более, 6-е маховые значительно короче 3-го и 4-го. Таким образом, получается впечатление, что крылья сороки менее узко приспособлены к полёту среди древесных зарослей, чем у сойки, но зато в какой-то степени допускают более быстрый полёт, а также перелёты на более дальние расстояния. Впрочем, у сороки внутривидовая изменчивость значительна; в частности, и летательный аппарат изменяется достаточно сильно. Так, например, у североафриканской формы P. p. mauretanica (Malherbe, 1845) крылья заметно короче, а хвост длиннее, чем у наших подвидов, что заставляет предполагать большую привязанность этой формы к лесу. С другой стороны, центральноазиатский подвид P. p. bottanensis (Delessert, 1840) имеет значительно более длинные крылья и более короткий хвост. Он населяет высокогорье, в частности субальпийскую зону. В этих местах сорока, перелетая через глубокие долины, совершает то и дело сравнительно дальние и высокие полёты.
Имеется ещё одна особенность крыла сороки, которая даёт возможность хотя бы слегка коснуться вопроса об его эволюции. У сороки характерным образом первое маховое рудиментарно, будучи совсем коротким, узким и заострённым. Однако в первом наряде это маховое значительно более крупное, т.е. нормальное для ворон. Таким образом, в течение онтогенетического развития сороки 1-е маховое сильно сокращается в размерах, теряя свою функцию и изменяя форму крыла. Как видно по всем приведённым рисункам, уменьшение 1-го махового у ворон должно делать крыло их более острым, удлинение его (до известных пределов) должно делать крыло более округлым.
Упомянутая здесь закономерность имеет общее значение. Известно, что в истории развития различных организмов имело место последовательное изменение специализации. Так, в различных группах птиц происходило приспособление то к быстрому полёту в открытых пространствах, то к маневренному полёту среди леса. Путь превращения острого крыла в тупое имеется лишь один: 1-е маховое, а за ним и 2-е сокращаются в длине; в случае необходимости в сильном изменении конфигурации крыла уменьшается длина и нескольких последующих маховых. Превращение же тупого крыла в острое теоретически можно себе представить двояким образом — путём удлинения 1-го махового, покуда оно не станет самым длинным, или же путём полной ликвидации 1-го махового. Практически, видимо, эволюция происходит только
Путь превращения острого крыла в тупое имеется лишь один: 1-е маховое, а за ним и 2-е сокращаются в длине; в случае необходимости в сильном изменении конфигурации крыла уменьшается длина и нескольких последующих маховых. Превращение же тупого крыла в острое теоретически можно себе представить двояким образом — путём удлинения 1-го махового, покуда оно не станет самым длинным, или же путём полной ликвидации 1-го махового. Практически, видимо, эволюция происходит только
вторым путём. Действительно, если представить себе, что 1-е маховое не длиннее половины 2-го, как это обычно бывает в тупом крыле, то незначительное увеличение его длины не сделает крыло более острым, скорее наоборот. Следовательно, в данном направлении естественный отбор неприложим. Дальнейшее же сокращение размеров 1-го махового одновременно с удлинением 2-го сразу же меняет форму крыла в смысле уменьшения округлости её вершины. Второе маховое, удлиняясь, приближается своей вершиной к вершине крыла и делает его более острым.
Случаев такого изменения вершины крыла имеется достаточно. Так, например, у наших щурок Merops крыло очень острое, 1-е маховое рудиментарно, 2-е — самое длинное. У близко родственной, так называемой ночной щурки Nyctiornis (2 вида), населяющей тропические леса, 1-е маховое развито нормально, крыло тупое. Не подлежит сомнению, что ночная щурка по форме крыла более близка к исходному типу, из которого настоящие щурки развились, приспособившись к длительному полёту в открытых местах с целью ловли летающих насекомых. Нужно полагать, что таким же образом утратили 1-е маховое вьюрки, приспособленные большей частью к стремительному полёту. В пределах славковых 1-е маховое может считаться индикатором формы крыла. Так, у сверчков Locustella 1-е маховое рудиментарно и крыло острое, у близкого рода Lusciniola 1-е маховое большое и крыло тупое. У пеночки-теньковки Phylloscopus collybita 1-е маховое длинное и крыло сильно закруглённое, у пеночки-веснички Ph.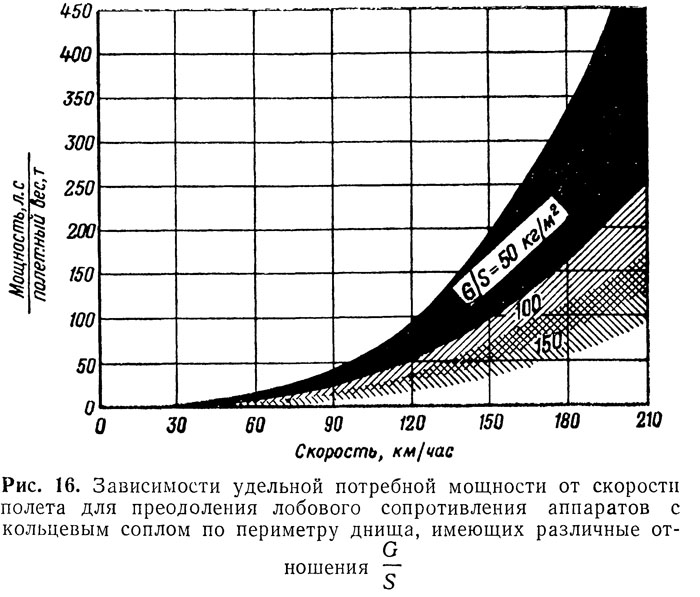 trochilus 1-е маховое маленькое и крыло более острое. Наконец, у пеночки-трещотки Ph. sibilatrix 1-е маховое рудиментарно и крыло наиболее острое. Таких примеров можно привести много, и все они свидетельствуют о том, что рудиментарное 1-е маховое (будь оно при наличии 9 или 10 развитых маховых) свидетельствует о происшедшем изменении в форме крыла.
trochilus 1-е маховое маленькое и крыло более острое. Наконец, у пеночки-трещотки Ph. sibilatrix 1-е маховое рудиментарно и крыло наиболее острое. Таких примеров можно привести много, и все они свидетельствуют о том, что рудиментарное 1-е маховое (будь оно при наличии 9 или 10 развитых маховых) свидетельствует о происшедшем изменении в форме крыла.
Из сказанного следует, что и у сороки произошло в недавнем прошлом изменение формы крыла — заострение вершины. Продолжается ли данный процесс и по сей день — сказать трудно. Возможно, что сорока, приспособившаяся в былое время к полёту в чаще, вторично вышла из леса на опушки и в перелески и современный тип ее полёта вполне адаптирован к её нынешнему образу жизни. Но, конечно, у нашей сороки возможно в будущем и дальнейшее приспособление к открытому ландшафту, как мы это видим у центральноазиатского подвида.
Другой своеобразный случай приспособления вороновой птицы к жизни в лесу являет собой кедровка Nucifraga caryocatactes. При сравнении с настоящими воронами сразу же бросается в глаза незначительная длина и большая ширина крыльев этой птицы (рис. 7). Од-
При сравнении с настоящими воронами сразу же бросается в глаза незначительная длина и большая ширина крыльев этой птицы (рис. 7). Од-
нако, по сравнению с сойкой, у кедровки крылья явно длиннее и уже. Размах у нее в 5.88 раза превосходит длину корпуса, ширина крыльев составляет 144.9% длины корпуса. Таким образом, у кедровок крылья ещё немного длиннее и уже, чем у сороки (табл. 1).
Таблица 1. Морфологические характеристики летательного аппарата
вороновых птиц
Вид Отношение размаха к длине корпуса Ширина крыла в % от длины корпуса Длина хвоста в % от длины корпуса Вес летательной мускулатуры в % от веса тела Вес подъёмной мускулатуры в % от веса опускающей мускулатуры Вес сердца в %о от веса тела
C. corax 7.36 123.4 134.6 15. 76 18.76 12.1
76 18.76 12.1
C. тгп 'к 6.94 142.8 132.1 16.54 16.85 10.6
C. frugilegus 7.08 134.6 128.1 18.52 14.23 11.96
C. monedula 6.53 125.8 118.7 18.61 16.76 13.27
P. pica 5.76 145.7 241.9 17.7 20.38 11.78
N. caryocatactes 5.88 144.9 143.5 15.43 22.25 1 1 .22
G. glandarius 5.65 154.3 157.5 17.41 22.8 11.09
P. pyrrhocorax 7.35 143.8 143.8 14.04 16.31 10.44
P. graculus 7.18 158.6 165.7 13.4 17.6 9.4
Наблюдения в природе говорят о том, что кедровка отлично приспособлена к жизни в лесу и ловко летает среди древесной чащи. Но она не избегает и перелётов через открытые пространства, поднимается выше леса и иногда играет в воздухе, чего сорока, например, никогда не делает. Автору приходилось наблюдать, как кедровки под осень стаями носились над лесом и уже после наступления темноты с шумом пикировали к месту ночлега.
Автору приходилось наблюдать, как кедровки под осень стаями носились над лесом и уже после наступления темноты с шумом пикировали к месту ночлега.
Из сказанного следует, что кедровка не утратила способности к дальним перелётам. Питаясь в значительной степени семенами хвойных, она в меньшей степени, чем сойка и кукша, нуждается в быстром передвижении среди лесной чащи, но зато должна быть в состоянии в поисках урожайных угодий перелетать из одного леса в другой, иногда на довольно большие расстояния. Эта особенность типа полёта кедровки хорошо выражена и в морфологии её крыла.
Что касается развития летательной мускулатуры, то оно у вороновых значительно более константно, чем внешняя морфология летательного аппарата. Вес летательной мускулатуры составляет у сойки 17.41%, у сороки 17.7%, у кедровки 15.43% от веса тела. Эти отношения находятся в пределах того, что мы видали у настоящих ворон.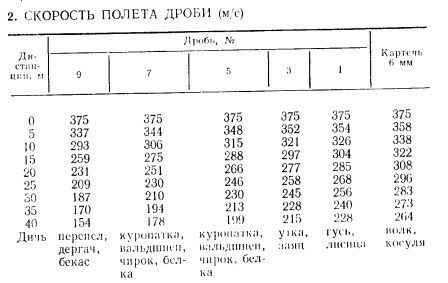 Более заметна разница в отношении между опускающей и подъёмной мускулатурой. У сойки вес подъёмной мускулатуры составляет 22.8%, у сороки 20.38%, у кедровки 22.25% веса опускающей мускулатуры, между тем как у настоящих ворон он всегда меньше 20% (табл. 1). От-
Более заметна разница в отношении между опускающей и подъёмной мускулатурой. У сойки вес подъёмной мускулатуры составляет 22.8%, у сороки 20.38%, у кедровки 22.25% веса опускающей мускулатуры, между тем как у настоящих ворон он всегда меньше 20% (табл. 1). От-
носительно большая мощность подъёмной мускулатуры у лесных вороновых находится в прямой связи с их образом жизни. Поминутные взлёты и посадки, при которых применяется трепещущий полёт, требуют большого напряжения подъёмной мускулатуры крыла, которая у настоящих ворон работает реже и менее интенсивно.
Сердце у лесных ворон достаточно большое; вес его составляет у сойки 11.09%, у сороки 11.78%, у кедровки 11.22% от веса тела. Видимо, меньшая продолжительность полёта у лесных видов компенсируется частыми взлётами, требующими большой затраты энергии.
Рис. 8. Силуэт летящего фазана Phasianus colchicus, вписанный в силуэт летящей кедровки Nucifraga caryocatactes
Небезынтересно сравнение лесных ворон с другой группой птиц, обладающих высоко маневренным полётом, а именно — с лесными куриными птицами.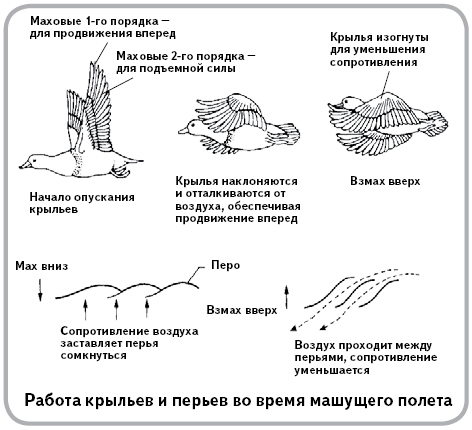 На рисунке 8 изображен силуэт летящей кедровки, в который вписан летящий фазан, приведённый к одинаковой с первой птицей длине корпуса. По сравнению с фазаном, кедровка кажется чрезвычайно крупнокрылой, и лишь пушистость оперения кедровки, её большая голова и длинный хвост в обычной обстановке скрадывают это впечатление. Разница же в величине крыльев объясняется в корне различной техникой полёта этих птиц. Маневренность полёта фазана является результатом мощного развития его летательной мускулатуры, позволяющей ему путём трепещущего полёта то резко затормозить движение, то быстро набрать скорость или рывком сменить направление полёта. По понятным причинам этот способ полёта очень неэкономен и может быть назван для куриных удобным только потому, что эти птицы вообще мало летают. В отличие от куриных птиц, лесные вороны постоянно перелетают с ветки на ветку, с дерева на дерево, и для них экономность полёта не менее важна, чем его маневренность. Но экономность полёта могла быть достигнута только путём
На рисунке 8 изображен силуэт летящей кедровки, в который вписан летящий фазан, приведённый к одинаковой с первой птицей длине корпуса. По сравнению с фазаном, кедровка кажется чрезвычайно крупнокрылой, и лишь пушистость оперения кедровки, её большая голова и длинный хвост в обычной обстановке скрадывают это впечатление. Разница же в величине крыльев объясняется в корне различной техникой полёта этих птиц. Маневренность полёта фазана является результатом мощного развития его летательной мускулатуры, позволяющей ему путём трепещущего полёта то резко затормозить движение, то быстро набрать скорость или рывком сменить направление полёта. По понятным причинам этот способ полёта очень неэкономен и может быть назван для куриных удобным только потому, что эти птицы вообще мало летают. В отличие от куриных птиц, лесные вороны постоянно перелетают с ветки на ветку, с дерева на дерево, и для них экономность полёта не менее важна, чем его маневренность. Но экономность полёта могла быть достигнута только путём
развития способностей к планированию, для чего необходимы большие крылья.
Сравнивая настоящих и лесных ворон, мы находим еще одно различие между ними, имеющее важное адаптивное значение. У настоящих ворон покровное оперение довольно плотно прилегает к телу, относительно мало распушено и гладкое, чем, несомненно, повышается обтекаемость тела, необходимая для их сравнительно быстрого полёта. Эта особенность достигается в ущерб термоизолирующим качествам пера. У лесных ворон, обладающих более медленным полётом, крайняя обтекаемость форм не является такой первоочередной проблемой, как у первой группы; это качество замещено у них более сильным развитием термоизолирующих свойств оперения. Действительно, у лесных ворон оперение очень пушистое и менее прилегает к телу, чем у настоящих ворон. Так, например, у сороки средняя ширина тела составляет 82 мм, у равной по величине галки — 67 мм. Сойка имеет ещё более пушистое оперение, чем сорока. Данная закономерность имеет широкое распространение среди птиц. Стоит лишь вспомнить лесных синиц, напоминающих пуховые шарики со вставленными спичками-хвостами, и тоненьких, зализанных ласточек.
В заключение рассмотрим своеобразную группу горных ворон, представленную всего двумя видами — клушицей РуттНосотах руттНо-согах и альпийской галкой РуттНосотах gтaculus. На первый взгляд эти птицы производят впечатление большой близости к настоящим воронам, но более тщательное изучение их не подтвёрждает такого предположения. Это небольшие вороновые птицы, чёрные, с ярко окрашенным клювом и ногами, населяющие в основном открытый ландшафт высокогорья.
Картина полёта клушицы очень своеобразна (рис. 9). Бросается в глаза большая длина крыльев - размах превышает длину корпуса в 7.36 раза, что соответствует относительной величине размаха у ворона. Однако ширина крыльев значительно больше, чем у ворона, подходя к таковой у вороны, так как она составляет 143.73% длины корпуса. Всё-таки удлинение крыла остаётся значительным.
Далее, для клушицы характерно особенно сильное развитие про-пеллирующих маховых. Число их равно 5, как у вороны, пропелли-рующие части их очень длинные, составляя более одной трети распростёртого крыла. 3-е, 4-е и 5-е маховые почти одинаковой длины; при большой длине крыла это производит очень своеобразное впечатление. Маховые, в частности и пропеллирующие, отличаются большой мягкостью, что является следствием малой удельной нагрузки на крылья. Хвост прямо срезанный и кажется коротким благодаря большому размаху крыльев. На самом деле его длина составляет 143.8% длины корпуса, превышая длину хвоста всех настоящих ворон
Число их равно 5, как у вороны, пропелли-рующие части их очень длинные, составляя более одной трети распростёртого крыла. 3-е, 4-е и 5-е маховые почти одинаковой длины; при большой длине крыла это производит очень своеобразное впечатление. Маховые, в частности и пропеллирующие, отличаются большой мягкостью, что является следствием малой удельной нагрузки на крылья. Хвост прямо срезанный и кажется коротким благодаря большому размаху крыльев. На самом деле его длина составляет 143.8% длины корпуса, превышая длину хвоста всех настоящих ворон
В соответствии с вышеописанными особенностями летательного аппарата клушица летает очень легко, сравнительно медленно взмахивая громадными крыльями, что особенно бросается в глаза при её небольших размерах — вес клушицы лишь немногим превышает вес обыкновенной галки. Она способна быстро пикировать и часто играет
Рис. 9. Клушица Pyrrhocorax pyrrhocorax.
Рис. 10. Альпийская галка Pyrrhocorax graculus.
в воздухе, то медленно рея, широко расставив пропеллирующие маховые, то стремительно скользя с подобранными крыльями, то попеременно пикируя и взмывая. В связи с большой величиной крыльев она хорошо парит, отлично используя восходящие воздушные потоки, характерные для горных стран. С сильными порывами ветра она легко справляется, максимально подбирая крылья и развивая таким образом, несмотря на свой малый вес и большие крылья, очень значительную скорость скольжения.
Альпийская галка почти тех же размеров, что клушица, но картина полёта у неё существенно иная (рис. 10). Размах крыльев у этой птицы превышает длину корпуса в 7.18 раза, т.е. меньше, чем у клушицы и
лишь немногим больше, чем у грача. Но ширина крыла составляет 158.61% длины корпуса, что заметно больше, чем у всех наших вороновых, даже лесных. Вершина крыла значительно суженная и более острая, чем у клушицы, так как 3-е маховое заметно длиннее 4-го и, тем более, 5-го махового. Пропеллирующих маховых на одно меньше, чем у клушицы, т.е. 4. Длина пропеллирующих частей маховых значительно меньше, примерно как у галки. Следовательно, крыло относительно компактное, в связи с чем стоит большая ширина маховых, особенно первостепенных,— признак, по которому альпийская галка тоже резко отличается от клушицы. Хвост, длинный, закруглённый, составляет 165.7% длины корпуса. Такая длина хвоста значительно больше, чем у всех настоящих ворон, больше даже, чем у сойки, и как будто не оправдывается особенностями образа жизни альпийской галки среди открытого, к тому же ещё высокогорного ландшафта.
Между тем альпийская галка населяет наиболее высокие части гор, у самых ледников и вечного снега. Самой характерной чертой её полёта является исключительная способность к парению. Часто можно видеть, как альпийская галка, взлетев с некрутого склона, после двухтрёх взмахов быстро и уверенно поднимается ввысь на неподвижно распростёртых крыльях. Играя в полуденное время, стаи этих птиц с непостижимой быстротой возносятся выше горных вершин, сразу пикируют, через минуту уже опять парят на громадной высоте и проделывают эти и подобные эволюции с лёгкостью, недоступной для других птиц. В случае необходимости альпийская галка летает активным полётом, довольно вяло взмахивая крыльями, но при первой возможности переходит на парение.
Борясь с сильными порывами ветра, альпийская галька, как и клушица, почти полностью прижимает крылья к телу, но для неё характерен ещё один особый приём. При подтянутых крыльях, когда вершинная часть их расположена параллельно телу, центр давления крыла передвигается далеко назад, и птица переходит в пике. Чтобы при максимально подобранных крыльях не пикировать, альпийская галка подгибает кисти настолько, что вершины крыльев складываются над хвостом и, таким образом, теряют свою несущую силу. В результате оттопыренной от тела остается только передняя часть крыла, и птица скользит с большой быстротой, однако не пикирует. Альпийская галка, несмотря на свою лёгкость, непомерно широкие крылья и длинный хвост, совсем не боится сильных порывов ветра и то возносится ими, то с легкостью противостоит им.
Исходя из того, что у вышеописанных двух видов морфология летательного аппарата неодинакова и техника полёта различна, можно предположить, что и летательная мускулатура у них развита неоди-
наково. Действительно, и в этом отношении между этими двумя видами заметны различия, что наглядно видно в таблице 2.
Таблица 1. Морфологические характеристики летательного аппарата клушицы Pyrrhocorax pyrrhocorax и альпийской галки Pyrrhocorax graculus
Вид Вес летательной мускулатуры в % от веса тела Вес подъёмной мускулатуры в % от веса опускающей мускулатуры Вес сердца в %о от веса тела
тіп тах Среднее тіп тах Среднее тіп тах Среднее
Р. pyrrhocorax 13.79 14.3 14.04 16.21 16.41 16.31 10.46 10.42 10.44
Р. graculus 12 14.5 13.4 15.05 20 17.6 5.5 11.8 9.4
У клушицы относительный вес летательной мускулатуры меньше, чем у всех ранее перечисленных вороновых птиц. Вероятно, это связано с тем, что данный вид хотя и много летает, но довольно много парит. У альпийской галки вес летательной мускулатуры ещё меньше в соответствии с тем, что она значительно больше парит и активным полётом пользуется немного. По той же причине и вес сердца у альпийской галки меньше, чем у клушицы и всех прочих вороновых птиц. Вообще, имеется основание полагать, что жизнь в разреженной атмосфере больших высот предъявляет высокие требования к работе сердца, вследствие чего у высокогорных животных сердце, как правило, крупнее, чем у видов, живущих в низменностях. Однако степень рабочей нагрузки на сердце в зависимости от типа полёта у птиц нарушает указанную закономерность. В результате у альпийской галки, населяющей самые большие высоты, но мало напрягающей при полёте свой мускульный аппарат, сердце сравнительно небольшое.
Заканчивая обзор лётных способностей наших вороновых птиц, остановимся кратко на оценке величины диапазона различий в этих способностях. В пределах монолитной группы вороновых мы видим приспособления и к дальним перелётам, даже скоростным (галка, ворон), и к маневренному полёту среди лесной чащи (сойка, кукша и др.). Качества эти, глубоко различные и взаимно исключающие друг друга, развились из одного типа крыла, большого, широкого и закруглённого. Активный полёт на далёкие расстояния развился в дальнейшем, как уже было указано, в направлении увеличения скорости в ущерб экономичности, но, с другой стороны, привел к специализации в виде парения, которое уже играет известную роль в полёте ворона и клушицы и стало основным способом передвижения у альпийской галки.
Особенности стрельбы
Скорости движения охотничьих целей
Таблица 1.
Быстрый полет птиц | 20 м/с (72 км/ч и более) |
Средняя скорость полета птиц | 15 м/с (54 км/ч) |
Быстрый бег зайца | 10-12 м/с (36-45 км/ч) |
Нормальный бег зайца | 7-8 м/с (25-28 км/ч) |
Величина перемещения охотничьих целей за время полета дроби (м)
Таблица 2.
Скорости движения целей (м/с) | Перемещения | |||
Дистанции (м) | ||||
20 | 30 | 40 | 50 | |
7 м/с Медленный бег зайца (дробь №1) | 0,42 | 0,63 | 0,98 | 1,26 |
10 м/с Быстрый бег зайца или медленный полет птицы (дробь №1) | 0,60 | 1,00 | 1,40 | 1,80 |
15 м/с Средняя скорость охотничьих птиц (дробь №5) | 0,90 | 1,50 | 2,25 | 3,00 |
Величина поправок на упреждение в корпусах дичи при угле встречи
заряда c целью 60-90°, т.е при выстреле по боковой цели и над головой
Таблица 3.
Величина поправок на упреждение в зависимости от угла встречи дробового заряда с целью
Таблица 4.
Точки прицеливания по неподвижным и движущимся целям
Таблица 5.
Самые быстрые птицы - Топ 10
Вопрос о том, какая из птиц самая быстрая в мире, всегда волновал человечество. Также как вопрос о том, какое самое быстрое млекопитающее на суше доминирует в воде и т. Д.
Какая самая быстрая птица в мире?Большинство наших читателей наверняка знают ответ на этот вопрос и знают, что это: сапсан ( Falco peregrinus ). Ответ на вопрос о втором и третьем месте известен также людям, которые уже изучили сводку самых быстрых животных в мире.Сегодняшнее заявление дает представление о том, какие птицы занимают следующие позиции. Интересно, что в заявлении также фигурирует самая большая птица в мире, которая хоть и не может летать, но развивает впечатляющую скорость.
Прежде чем мы перейдем к таблице рекордов скорости птиц, краткий обзор самых быстрых представителей птичьего семейства.
Самые быстрые птицы в мире - характеристика Golden Eagle ( Aquila chrysaetos ). 1. Сапсан ( Falco peregrinus ) - самая быстрая птица в миреСамая быстрая птица представлена соколом-сапсаном.Во время пикировочного полета сокол-сапсан может развивать скорость до 350 км / час! Рекордная скорость сокола, которую удалось измерить, составила 389 км / ч (242 мили в час) .
Сапсан ( Falco peregrinus ) - первое место. 2. Золотой орел ( Aquila chrysaetos )Некоторые источники утверждают, что во время пикирования сокол может развивать скорость более 400 км / ч (248 миль / ч)…
Второе место занял Беркут.Хотя эта птица намного крупнее сокола и не может похвастаться своей маневренностью, во время пикирования достигает скорости до 320 км / ч (199 миль / ч).
Golden Eagle ( Aquila chrysaetos ) - второе место. 6. Стриж обыкновенный ( Apus apus )Известны его действия во время ухаживания, когда орел хватает камень с земли, поднимает его и бросает с луком, затем ныряет с головокружительной скоростью, ловит его на лету, прежде чем камень успевает коснуться земли. …
Стриж - удивительная птица - большую часть жизни он проводит в воздухе.Он охотится и ест в воздухе, в воздухе даже спит ... Природа настолько в высшей степени приспособила стрижа к полету, что его ноги были укорочены, и у птицы проблемы с отрывом от земли. Поэтому стрижи в основном выбирают высокие здания, дымоходы или вертикальные поверхности, с которых он может легко начать скользить.
Common Swift ( Apus apus ) - четвертое место. Самые быстрые птицы в мире - ТОП 10Хотя может показаться, что он похож на ласточку, на самом деле это не так. Это от нее во много раз быстрее. Common Swift развивает максимальную скорость 170 км / ч (106 миль в час), хотя некоторые источники утверждают, что он может летать со скоростью 180 км / ч (112 миль в час) в горизонтальном полете.
- Сапсан ( Falco peregrinus ) - 389 км / ч (242 миль / ч) / ныряние
- Golden Eagle ( Aquila chrysaetos ) - 320 км / ч (199 миль / ч) / дайвинг
- Степной орел ( Aquila nipalensis ) - 300 км / ч (186 миль / ч) / дайвинг
- Кречет ( Falco rusticolus ) - 209 км / ч (130 миль / ч) / дайвинг
- Краснохвостый ястреб ( Buteo jamaicensis ) - 190 км / ч (118 миль / ч) / дайвинг
- Common Swift ( Apus apus ) - 171 км / ч (106 миль / ч)
- Игольчатый хвост с белым горлом ( Hirundapus caudacutus ) - 170 км / ч (105 миль / ч)
- Eurasian Hobby ( Falco subbuteo ) - 160 км / (100 миль / ч)
- Frigatebird - 153 км / ч (95 миль / ч)
- Spur-winged Goose ( Plectropterus gambensis ) - 142 км / ч (88 миль / ч)
- Домашний голубь ( Columba) ) - 140 км / ч (87 миль / ч)
- Красногрудый Merganser ( Mergus serrator ) - 130 км / ч (81 миль / ч)
- Белозубый Swift ( Apus caffer ) ) - 124 км / ч (77 миль / ч)
- Canvasback ( Aythya valisineria ) - 116 км / ч (73 миль / ч)
- Eider duck ( Somateria mollissima ) - 113 км / ч (70 миль / ч)
- Common Teal ( Anas crecca ) - 109 км / ч (69 миль / ч)
- Странствующий альбатрос ( Diomedea exulans ) - 108 км / ч (67 миль / ч)
- Wild Duck ( Anas pla tyrhynchos ) - 105 км / ч (65 миль / ч)
- Northern Pintail ( Anas acuta ) - 105 км / ч (65 миль / ч)
- Кряква / дикая утка ( Anas platyrhynchos ) -105 км / ч (65 миль / ч)
- Оушан - 100 км / ч (63 миль / ч)
- Колибри Анны (Calypte anna) - 98 км / ч (61 миль / ч)
- California condor ( Gymnogyps californianus ) -90 км / ч (56 миль / ч)
- Страус обыкновенный ( Struthio camelus ) -72 км / ч (45 миль / ч)
В позиции 24 находится Common Ostrich , который развивает максимальную скорость при беге 72 км / ч (45 миль / ч).Мы познакомились с источником, сообщающим, что на очень коротком расстоянии птица может бегать со скоростью 98 км / ч (61 миль / ч). Поскольку, однако, мы не можем подтвердить эту скорость, мы представляем 72 км / ч (45 миль / ч).
Самые быстрые птицы в мире - 10 самых быстрых птиц на диаграмме [км / ч]Стоит также обратить внимание на то, сколько позиций в списке занимают птицы семейства уток - Anatidae .
Самые быстрые птицы ТОП 10 - скорость на диаграмме в км / ч Самые быстрые птицы в мире - 10 вершин скорости на диаграмме
[миль / ч] TOP 10 самых быстрых птиц - диаграмма скорости в миль / ч. Рекомендуемые
Скорости полета среди видов птиц: аллометрические и филогенетические эффекты
Abstract
Ожидается, что скорость полета будет увеличиваться с увеличением массы и нагрузки на крыло у летающих животных и самолетов по фундаментальным аэродинамическим причинам. Предполагая геометрическое и динамическое сходство, прогнозируется, что крейсерская скорость полета будет варьироваться как (масса тела) 1/6 и (нагрузка на крыло) 1/2 среди видов птиц. Чтобы проверить эти правила масштабирования и общую важность массы и нагрузки на крыло для скорости полета птиц, мы использовали радар слежения для измерения скорости взмахов крыльев отдельных особей или стай перелетных птиц, визуально идентифицируемых по видам, а также их высоты и ветра на высотах птицы летели.Эквивалентные воздушные скорости (скорректированные с учетом плотности воздуха на уровне моря, U e ) 138 видов, массой 0,01–10 кг, были проанализированы с точки зрения биометрии и филогении. Показатели масштабирования по отношению к массе и нагрузке на крыло были значительно меньше, чем прогнозировалось (около 0,12 и 0,32, соответственно, с аналогичными результатами для анализов, основанных на видах и независимых филогенетических контрастах). Эти низкие показатели масштабирования могут быть результатом эволюционных ограничений на диапазон скоростей полета птиц, противодействующих слишком низкой скорости полета среди видов с низкой нагрузкой на крыло и слишком высокой скорости среди видов с высокой нагрузкой на крыло.Такое сжатие диапазона скоростей частично достигается за счет геометрических различий, при этом удлинение показывает положительную взаимосвязь с массой тела и нагрузкой на крыло, но требуются дополнительные факторы, чтобы полностью объяснить небольшой масштабный показатель U e в отношении нагрузки на крыло. . Кроме того, масса и нагрузка на крыло составляли лишь ограниченную часть вариации в U e . Филогения в сочетании с нагрузкой на крыло была мощным фактором, объясняющим вариации в U, , и .Эти результаты демонстрируют, что функциональные летные адаптации и ограничения, связанные с различными эволюционными линиями, имеют важное влияние на крейсерскую скорость взмахивания крыльев, которая выходит за рамки общих аэродинамических эффектов масштабирования массы и нагрузки на крыло.
Сведения об авторе
Анализ изменения скорости полета среди видов птиц важен для понимания полета. Мы проверили, соответствует ли крейсерская скорость разных видов перелетных птиц в машущих полетных весах с учетом массы тела и нагрузки крыльев предсказаниями аэродинамической теории, и в какой степени филогения дает дополнительное объяснение вариации скорости.Скорость полета измерялась с помощью радара слежения за видами птиц размером от 0,01 кг (мелкие воробьиные) до 10 кг (лебеди). Эквивалентные воздушные скорости 138 видов варьировались от 8 до 23 м / с и не масштабировались так резко по отношению к массе и нагрузке на крыло, как прогнозировалось. Это говорит о том, что существуют эволюционные ограничения на диапазон скоростей полета, которые достигаются птицами, которые противодействуют слишком медленным и слишком высоким скоростям у видов птиц с низкой и высокой нагрузкой на крыло, соответственно. Помимо влияния размера тела и морфологии крыла на скорость полета, мы также показываем, что филогения составляла важную часть оставшейся вариации скорости между видами.Различия в летном аппарате и поведении видов разного эволюционного происхождения, с разной экологией и стилями полета, вероятно, во многом повлияют на крейсерские летные характеристики.
Образец цитирования: Алерстам Т., Розен М., Бэкман Дж., Эриксон П.Г.П., Хеллгрен О. (2007) Скорость полета среди видов птиц: аллометрические и филогенетические эффекты. PLoS Biol 5 (8): e197. https://doi.org/10.1371/journal.pbio.0050197
Академический редактор: Бен Шелдон, Оксфордский университет, Великобритания
Поступила: 2 февраля 2007 г .; Принята к печати: 16 мая 2007 г .; Опубликовано: 17 июля 2007 г.
Авторские права: © 2007 Alerstam et al.Это статья в открытом доступе, распространяемая в соответствии с условиями лицензии Creative Commons Attribution License, которая разрешает неограниченное использование, распространение и воспроизведение на любом носителе при условии указания автора и источника.
Финансирование: Эта работа финансировалась за счет грантов Шведского исследовательского совета по естественным наукам и Шведского исследовательского совета при TA. Реконструкция радара для отслеживания птиц финансировалась за счет грантов Фонда Кнута и Алисы Валленбергс и Шведского совета по планированию и координации исследований.
Конкурирующие интересы: Авторы заявили, что конкурирующих интересов не существует.
Сокращения: АПК, Информационный критерий Акаике; df, степени свободы
Введение
Согласно фундаментальной аэродинамике подъемная сила ( L ), создаваемая на крыле, связана со скоростью полета ( U ) следующим образом: где ρ - плотность воздуха, S - площадь крыла, а C L - коэффициент подъемной силы [1–3].В горизонтальном крейсерском полете L уравновешивает вес ( м × g ), и ожидается, что самолет, а также животные будут летать со значением C L или близким к нему, обеспечивая максимально эффективное аэродинамическое сопротивление. При условии, что это значение C L примерно одинаково для разных видов птиц (что требуется для динамического сходства) [1], из этого следует, что крейсерская скорость полета среди видов птиц, как ожидается, будет зависеть от массы тела и нагрузки на крыло ( Q = м × g / S ) как U ∝ м 1/6 и U ∝ Q 1/2 , соответственно (с прежней пропорциональностью, основанной также на предположении геометрического подобия; i.е., S варьируется от м 2/3 ). Эти правила масштабирования также использовались для сравнения общих скоростей широкого круга летчиков, от самых маленьких насекомых до самых больших самолетов [1,4–6].
В связи с отсутствием надежных измерений воздушной скорости различных видов птиц в крейсерском (миграционном) полете на большие расстояния, для исследования этих правил масштабирования использовались теоретически полученные скорости полета для видов разной массы и морфологии крыльев [4,5,7 –10].Отклонения от ожидаемого показателя масштабирования по отношению к массе были обнаружены из-за отклонений от геометрического сходства - более крупные птицы часто имеют пропорционально большую площадь крыльев и размах [2,5,9–11]. Помимо отклонения от геометрического подобия, существуют дополнительные возможные причины отклонения скорости полета птиц от правил аэродинамического масштабирования. Адаптация к полету, связанная с экологией и филогенией птиц, может иметь последствия для их крейсерской скорости полета, а различные режимы полета (непрерывный или прерывистый взмах) могут ограничивать скорость птиц [2,10].
Полная оценка применимости правил аэродинамического масштабирования должна основываться не на теоретически выведенных скоростях, а на эмпирических измерениях воздушных скоростей большого количества видов птиц в естественных крейсерских полетах. Здесь мы представляем отслеживающие радиолокационные измерения скорости полета 138 видов из шести основных монофилетических групп [12], которые были проанализированы в связи с биометрией (m, S, и размах крыльев b) и эволюционным происхождением (что отражено филогенетическими данными). группа).Все скорости, указанные здесь, относятся к взмахам крыльев при крейсерской скорости перелетных птиц. Ограничивая анализ миграционным полетом, мы ожидаем, что птицы будут летать с воздушной скоростью, близкой к скорости, связанной с максимальной подъемной силой и сопротивлением [13]. Все скорости обозначают эквивалентные воздушные скорости ( U e ) с поправкой на плотность воздуха на уровне моря [14,15].
РезультатыВзаимосвязи между U e и m и Q для всех различных видов представлены на рисунке 1, а линии показывают аллометрические отношения в соответствии с сокращенными регрессиями по большой оси (таблица 1).Средняя скорость полета 138 особей колебалась от 8 до 23 м / с. Хищные птицы, певчие птицы, стрижи, чайки, крачки и цапли имели скорость полета в нижней части этого диапазона, в то время как голуби, некоторые кулики, ныряльщики, лебеди, гуси и утки летали быстро в диапазоне 15–20 РС. Бакланы, журавли и поморники были среди видов, летающих со средней скоростью, около 15 м / с. Ныряющие утки достигли самых высоких средних скоростей в нашей выборке, у некоторых видов они превышали 20 м / с, вплоть до 23 м / с (протокол S1).
Рисунок 1. Скорости полета птиц ( U e ; м / с) в зависимости от массы тела (кг) и нагрузки на крыло (Н / м 2 ) для 138 видов шести основных монофилетических групп
Линии показывают масштабные отношения U e = 15,9 × (масса) 0,13 и U e = 4,3 × (нагрузка на крыло) 0,31 , рассчитанные с помощью сокращенной регрессии по большой оси для всех видов (Таблица 1 ). Все оси в логарифмическом масштабе.На вставках показаны средние значения (± стандартное отклонение) для шести основных филогенетических групп по отношению к этим масштабным линиям. Виды одной и той же группы имеют тенденцию летать с одинаковой скоростью, и филогенетическая группа является важным фактором для объяснения различий в U e .
https://doi.org/10.1371/journal.pbio.0050197.g001
Анализ масштабирования на уровне видов устойчив к возможным ошибкам из-за нескольких следов для каждого вида и внутривидовых вариаций скорости (см. Материалы и методы и Таблица 1).Поскольку виды не представляют собой эволюционно независимую точку данных, мы также рассчитали показатели масштабирования путем анализа независимых филогенетических контрастов [16] в соответствии с процедурой и филогенезом [12], представленными в Протоколе S2. Мы использовали хорошо разрешенную молекулярную филогению Ericson et al. [12] для наших филогенетических анализов и классификаций. Результаты масштабирования с поправкой на филогенетическую зависимость очень близко совпадали с показателями, рассчитанными на уровне видов (таблица 1), демонстрируя, что показатели масштабирования для U e по отношению к m , а также Q (0.12 и 0,32 соответственно; филогенетический контрастный анализ) были меньше прогнозируемых значений 0,17 и 0,50 соответственно. Для масштабирования U e по сравнению с m разница от прогнозируемого значения была на уровне значимости 0,05 для анализа филогенетических контрастов, и разница не была статистически значимой для выборки скоростей, скорректированных с учетом внутри- видовая изменчивость (таблица 1).
В рамках различных основных филогенетических групп (уровень видов), как определено в Протоколе S1 (см. Рисунок 1), коэффициенты масштабирования U e по отношению к m были значительно меньше, чем прогнозируемое значение 0.17 среди двух групп. Лебеди / гуси / утки показали замечательный отрицательный коэффициент масштабирования -0,15 (отличие от прогноза t = 13,40, степени свободы (df) = 25 и p <0,0001), а соколы / вороны / певчие птицы показали масштабирование показатель 0,08, который был явно меньше ожидаемого ( t = 6,01, df = 37 и p <0,0001). Для других четырех групп показатели масштабирования находились в диапазоне от 0,12 до 0,20 и существенно не отличались от прогнозируемого значения ( p > 0.2). Соответствующие показатели масштабирования U e по отношению к Q значительно отличались от прогнозируемого значения 0,5 среди трех групп, фламинго / голуби / стрижи (показатель 0,28, t = 3,22, df = 5, и p = 0,023), водолазы / бакланы / пеликаны / цапли / аисты / журавль (показатель степени 0,36, t = 2,59, df = 15 и p = 0,021) и соколы / вороны / певчие птицы (показатель 0,28 , t = 4.88, df = 37 и p <0.0001). Для остальных трех групп показатели масштабирования находились в диапазоне от 0,42 до 0,54 и существенно не отличались от прогнозируемого значения ( p > 0,4).
Чтобы определить, были ли геометрические различия в форме крыла, связанные с различиями в массе и нагрузке на крыло, мы исследовали, было ли значительно увеличено удлинение с м и Q . Соотношение сторон - это безразмерная мера формы крыла (= b 2 / S ).Мы обнаружили значительные отклонения от изометрии с положительным масштабированием соотношения сторон до м , а также Q ( p <0,01 на основе всех видов [ n = 129] и p <0,05 на основе независимые филогенетические контрасты [ n = 17] для обоих масштабных соотношений).
Мы также исследовали объяснительную силу m, Q, соотношение сторон и филогенетическую группу, чтобы учесть вариацию в U e (рис. 2).На массу приходилась лишь небольшая часть изменения скорости полета, в то время как, как и ожидалось, скорость была гораздо более тесно связана с нагрузкой на крыло. Между U e и соотношением сторон наблюдалась значительная положительная корреляция, но соотношение сторон не дало улучшения общих линейных моделей (на основе информационного критерия Акаике [AIC] [17]) в сочетании с Q или филогенетической группой.
Рис. 2. Объяснение изменения средней скорости полета ( U e ; м / с) среди видов птиц по различным комбинациям переменных и факторов
Объясняющая сила (скорректированная R 2 ) различных Общие линейные модели со значимыми независимыми переменными (***, p <0.001). Филогенетическая группа и нагрузка на крылья являются ключевыми факторами, объясняющими различия в скорости полета среди видов птиц. Были рассчитаны общие линейные модели для всех различных комбинаций массы тела, нагрузки на крыло, соотношения сторон и филогенетической группы, за исключением комбинаций, включающих как массу тела, так и нагрузку на крыло (из-за взаимозависимости между этими переменными). Сложные модели (включая комбинации переменных) представлены только в том случае, если AIC улучшен по сравнению с моделями, основанными на отдельных независимых переменных [17].Это относилось только к модели, включающей как филогенетическую группу, так и нагрузку на крыло. ΔAIC указывает разницу в оценке AIC от наиболее эффективной модели (при ΔAIC = 0). Статистика испытаний была следующей (в скобках) для модели, включая массу ( F 1,136 = 20,0, p <0,001), соотношение сторон ( F 1,127 = 28,6, p <0,001), крыло нагрузка ( F 1,127 = 122,6, p <0,001), филогенетическая группа ( F 5,132 = 34.5), и филогенетическая группа плюс нагрузка на крыло ( F 6,122 = 39,6, p <0,001).
https://doi.org/10.1371/journal.pbio.0050197.g002
Наиболее важным фактором для объяснения вариации в U e была филогенетическая группа; виды одной и той же группы имели тенденцию летать с одинаковой характеристической скоростью. Группы, включающие хищных птиц и цапель, имели в среднем низкие скорости полета в зависимости от их массы и нагрузки на крылья, в то время как средняя скорость для групп, включая певчих птиц и куликов, была выше общих линий шкалы (Рисунок 1).Одна только основная филогенетическая группа составляла значительную часть вариации в U e (скорректированная R 2 = 0,55), а общая линейная модель, включающая как Q , так и филогенетическую группу, была наиболее удовлетворительной моделью, согласно в AIC (с поправкой R 2 = 0,64; рисунок 2).
Наши оценки объяснения, обеспечиваемого филогенетическим компонентом, в соответствии с рисунком 2, вероятно, будут консервативными из-за широкой группировки по всей современной филогении птиц.Если использовались более узкие монофилетические группы на уровне семьи (20 филогенетических групп), филогенетическая группа составляла фракцию до 0,68 (скорректированный R 2 ; F 19,118 = 16,4 и p <0,001 ) вариации в U e , а для модели, включающей как филогенетическую группу, так и Q , эта доля увеличилась до 0,71 (скорректировано R 2 ; F 20,108 = 16.4, и p <0,001). Однако эти модели имели положительные значения ΔAIC (+8,1 и +28,8, соответственно) по сравнению с лучшей моделью на Рисунке 2 и, таким образом, были менее удовлетворительными при рассмотрении сочетания соответствия и сложности [17].
Обсуждение
В результате нашего анализа были получены два основных результата; (1) скорость полета у разных видов птиц значительно отличается от массы и нагрузки на крыло, чем предсказывалась на основе основных аэродинамических принципов, и (2) филогенетическая группа внесла очень важный вклад в объяснение значительного различия в скорости полета птиц, которое оставалось даже после учитывались биометрические размеры видов птиц.
Масштабирование скорости полета
Показатели масштабирования упали ниже прогнозируемых значений для обоих тестируемых соотношений, для U e по сравнению с м , а также для U e по сравнению с Q . Прогнозируемые показатели масштабирования были основаны на предположениях геометрического и динамического подобия. Могут ли отклонения от одного или обоих этих предположений объяснить наши результаты? Более ранние исследования показали, что виды птиц, в среднем, не идентичны геометрически, но более крупные виды обычно имеют пропорционально более длинный размах крыльев и большее соотношение сторон [2,5,10].Это было подтверждено для образца в настоящем исследовании со значительным положительным масштабированием соотношения сторон до м , а также Q .
Общий коэффициент масштабирования 0,14 для скорости полета по отношению к массе тела был рассчитан для теоретической скорости полета после учета небольшой положительной аллометрии в размере крыла для большой выборки видов птиц [9]. Это хорошо согласуется с соответствующей экспонентой для наблюдаемых скоростей в этом исследовании, что делает отклонение от геометрического подобия вероятным объяснением этого результата.Отрицательный показатель масштабирования U e по отношению к м для лебедей, гусей и уток может быть следствием уменьшения запаса мощности полета с увеличением размера, ограничивающим самые большие летуны, такие как лебеди, летать близко к минимуму. силовая скорость, а не более высокая скорость, связанная с максимальным эффективным аэродинамическим сопротивлением [18,19]. Такая ограниченная скорость полета для самых крупных пилотов также будет иметь эффект снижения общих показателей масштабирования, что дает еще одно дополнительное объяснение наблюдаемым результатам в этом исследовании.
Динамическое сходство отражается числом Рейнольдса, которое будет различаться для разных видов птиц пропорционально их размеру (длине) и скорости [20]. Число Рейнольдса показывает 15-кратный диапазон среди видов в нашей выборке (примерно от 25000 до 375000 на основе средней хорды крыла, S, / b, при измерении длины). Такой диапазон чисел Рейнольдса вполне может быть достаточно большим, чтобы вызвать значительные отклонения от динамического подобия. Основным ожидаемым последствием будет снижение коэффициента сопротивления трения для птиц с большим числом Рейнольдса (т.е., крупные и быстрые птицы), что приводит к повышенной оптимальной крейсерской скорости среди этих видов [14,20]. Таким образом, такое отклонение от динамического подобия, как ожидается, проявится как увеличенный показатель масштабирования для U e по сравнению с m (а также для U e по сравнению с Q ), а не как показатель масштабирования. ниже, чем ожидалось, как в этом анализе.
Принимая во внимание противоположное влияние на показатели масштабирования отклонений от геометрического и динамического сходства, соответственно [1], мы заключаем, что только отклонение от геометрического сходства может объяснить, почему показатель масштабирования для U e падает по сравнению с m значительно ниже одной шестой среди птиц, совершающих крейсерский перелет.
Являются ли геометрические различия достаточным объяснением также того факта, что показатель масштабирования для U e по сравнению с Q явно упал ниже ожидаемого значения в половину? Один из способов оценить это - вычислить коэффициент масштабирования для скорости полета в зависимости от нагрузки по размаху ( м × г / бар 2 , где b - размах крыла). Нагрузка на размах эквивалентна нагрузке на крыло, разделенной на удлинение, и для птиц, различающихся геометрическими формами крыльев, ожидается, что крейсерская скорость полета будет наиболее точно масштабироваться с квадратным корнем из нагрузки на размах (при геометрическом сходстве скорость полета прогнозируется масштабируемой с один и тот же показатель половины в зависимости от нагрузки на размах и нагрузки на крыло) [5].
Коэффициент масштабирования для U e в зависимости от нагрузки (уровень вида, показатель 0,36 с 95% доверительным интервалом 0,31–0,40, n = 129 и филогенетические контрасты, показатель 0,37 с доверительным интервалом 95% 0,26–0,48, n = 17) превысило это значение по сравнению с Q (с соответствующими показателями 0,31 и 0,32, соответственно, таблица 1), хотя все еще значительно ниже прогнозируемого значения в половину. Это предполагает, что геометрические различия объясняют часть, но не все, несоответствия между наблюдаемым и ожидаемым масштабированием U e по сравнению с Q .Отклонение от динамического подобия в его наиболее простой форме (что отражается в различиях в числе Рейнольдса) будет способствовать увеличению, а не уменьшению показателя масштабирования по отношению к прогнозируемому, и поэтому не может дать никаких полезных дополнительных объяснений в этом случае (см. Выше ). Тем не менее, динамические различия других видов могут существовать по причинам, которые, как известно, трудно предсказать для махового полета. Будущие исследования вихревых паттернов, связанных с взмахами полета разных видов, будут важны для демонстрации возможных динамических различий между видами (см. Ниже).
Мы предполагаем, что неожиданно маленький показатель масштабирования для U e по сравнению с Q может быть результатом общих эволюционных сил, действующих для увеличения крейсерских скоростей для видов с наименьшими нагрузками на крыло и снижения скоростей для видов с наибольшими нагрузками на крыло. . Виды птиц в нашем анализе показывают примерно 10-кратную разницу в диапазоне значений Q (примерно от 15 до 150 Н / м 2 , рис. 1). С наблюдаемым масштабным показателем скорости полета 0.31, этот диапазон Q связан с двукратной (10 0,31 = 2,0) разницей в скорости полета. Однако с прогнозируемым показателем масштабирования 0,5 мы ожидали бы более чем трехкратную разницу в крейсерской скорости (10 0,5 = 3,2). Учитывая, что птицы с низким значением Q (около 15 Н / м 2 ) летают со скоростью около 10 м / с (по наблюдениям), птицы с высоким Q (около 150 Н / м 2 ) будут летать. на скорости 32 м / с по общим правилам аэродинамического масштабирования.Это может быть непрактично быстрым и трудным для согласования с летными характеристиками в ситуациях старта, посадки, маневров стайкой и т. Д. И наоборот, учитывая, что птицы с высоким значением Q летают со скоростью около 20 м / с (по наблюдениям), виды с низким Q будет летать со скоростью всего около 6 м / с в соответствии с общими правилами аэродинамического масштабирования. Такие очень низкие скорости будут невыгодны из-за чувствительности к ветру, уязвимости для хищников и т. Д. Следовательно, кажется разумным ожидать, что существуют эволюционные силы, действующие, чтобы сузить диапазон крейсерских скоростей полета среди видов птиц [5] и тем самым уменьшить показатель масштабирования для U e по сравнению с Q .Такое сжатие диапазона скоростей полета частично достигается за счет общих геометрических различий между видами (более широкие соотношения аспектов между видами с большей массой и нагрузкой на крыло, как обсуждалось выше), но с дополнительными неизвестными механизмами, возможно, связанными с разной кинематикой полета или разными мышцами взаимодействие между видами, по-видимому, необходимо для полного объяснения ограниченного диапазона скоростей полета среди видов птиц.
Ограничивающий полет, кажется, является режимом для мелких птиц (в основном воробьиных), чтобы уменьшить затраты на быстрый полет [1,2,10,21], в то время как полет с закрылками, используемый многими хищными птицами, связан с сокращением крейсерского полета. скорость [21].Оба эти стиля прерывистого полета используются видами с низким или средним значением Q (Рисунок 1), и, оказывая противоположное влияние на скорость полета, они вряд ли могут дать достаточное объяснение низкой степени масштабирования U . e против Q среди видов птиц в целом.
Изменчивость скорости полета
Анализ размеров показал, что масштабные соотношения между нагрузкой на крыло и общей массой значительно различаются у разных типов птиц [5,10].Ожидаемым следствием этого является то, что нагрузка на крыло будет более надежным показателем скорости полета, объясняя больше вариаций скорости полета у разных видов птиц, чем массу тела [1,5]. Это ожидание полностью подтвердилось в настоящем исследовании: на Q приходится почти половина вариации U e между видами, тогда как на m объясняется только 12% этой вариации (рис. 2). Однако наши выводы о том, что Q по-прежнему оставил большую часть вариации скорости полета необъяснимой, и что филогенетическая группа составляла значительную часть этой оставшейся вариации, были неожиданными из более ранних анализов, основанных на теоретически рассчитанных скоростях полета [5,10].
Каковы причины разницы в скорости полета филогенетических групп? Различия в режимах полета и использование ограничивающего полета многими воробьиными были предложены в качестве объяснения важных групповых отклонений от аэродинамических прогнозов оптимальной скорости полета птиц [15]. Мы условно отнесли разные виды птиц к трем основным режимам полета, основанным на нашем собственном полевом опыте; (1) непрерывное хлопанье (например, куликов и уток), (2) прерывистое хлопанье с короткими фазами планирования (хищники, стрижи и ласточки) и (3) ограниченный полет (многие, но не все воробьиные используют этот режим прерывистого хлопанья с фазы складывания крыла). U e значительно различались между листовками в этих трех категориях ( p <0,001, скорректированный R 2 = 0,26 и F 2135 = 25,1), а также объясняющая сила модели, включающей оба режима полета и Q были высокими ( p <0,001, скорректированный R 2 = 0,60 и F 3,125 = 64,5). Это говорит о том, что различие в режимах полета является одним из элементов, влияющих на характерные крейсерские скорости полета филогенетических групп.
В зависимости от своего экологического образа жизни и кормления птицы приспосабливаются к различным аспектам летных характеристик, например, скорости, маневренности, подъемной силы, эвакуации, взлета, стоимости транспортировки и мощности [2,10]. Эти адаптации могут иметь последствия для летного аппарата (анатомии, физиологии и работы мышц) и поведения в полете, что может ограничивать крейсерскую скорость полета. Различия в соотношении мощности и скорости между разными видами [22] и в эффективности мышц (преобразование входной метаболической мощности в выходную механическую) с массой и скоростью полета [23,24] могут быть связаны с такими дифференциальными сложными адаптациями полета среди птицы.Ограничения скорости полета также могут быть связаны с различиями в гидродинамике и вихревых структурах, которые были исследованы только для нескольких видов [25–27]. Переменная воздушная скорость может быть связана с высоким КПД мощности, если сопровождается правильным изменением частоты и амплитуды гребков крыла [28,29].
Виды, летающие на сравнительно малых крейсерских скоростях, часто используют термическое парение (хищники и аисты), приспособлены для охоты и переноски грузов (хищные птицы) или для взлета и посадки в густой растительности (цапли).Связанные с этими летными привычками, у них более низкое отношение подъемной (supracoracoideus) к депрессорной (грудной) мышце полета (особенно низкое у хищных птиц) по сравнению с куликами и анатидами [2]. Мы предполагаем, что функциональные различия в летном аппарате и мускулатуре птиц, различающихся по жизни и стилю полета (различия, часто связанные с эволюционным происхождением), оказывают значительное влияние на производительность и скорость птиц при длительном крейсерском полете. Таким образом, наши результаты убедительно показывают, что существует разнообразие характеристик крейсерского полета у разных типов птиц помимо общих масштабных эффектов массы и нагрузки на крыло, которые еще предстоит изучить и понять, аэродинамически [30], кинематически [26,31] ], физиологически [22], а также экологически [2,10].
Материалы и методы
Сопровождение радиолокационных измерений.
Наш основной набор данных, основанный на измерениях с помощью радара слежения в Швеции и Арктике в 1979–1999 гг., Состоит из 1399 следов 102 идентифицированных видов со средним временем отслеживания 369 с (диапазон 20–2220 с). Высоты колебались от уровня моря до 3600 м. Количество следов для каждого вида варьировалось от одного до 240, среднее значение U e (с SD), вертикальная скорость, а также информация о количестве следов, времени следа и биометрических данных приведены для каждого вида в протоколе S1. .
Обширный дополнительный набор данных об эквивалентных скоростях полета идентифицированных птиц, полученных с помощью аналогичных радиолокационных методов слежения, был опубликован на основе работы Бруно Брудерера и его исследовательской группы в Швейцарии, Германии, Израиле и Испании [15]. В наш анализ были включены данные о скорости полета по следам птиц при естественном миграционном полете (за исключением выпущенных птиц и парящего полета). Этот дополнительный набор данных включал 64 вида, а с 28 видами, общими для двух наборов данных, объединенные данные в сумме составили 138 видов (Протокол S1).Средние значения U e для общих видов существенно не различались между двумя наборами (t-критерий парной выборки, t = 1,28 и p = 0,21), и мы использовали взвешенные (в соответствии с количеством треков) общее среднее значение U e для этих видов в нашем анализе.
Большая часть данных о скорости полета была измерена в 1979–1999 гг. С помощью радиолокационных исследований в пяти местах на юге Швеции и в ходе двух ледокольных экспедиций в Арктику (подробные методы см. В [19,32]).Цели идентифицировались по видам и размерам с помощью телескопов одновременно с регистрацией радаров, обеспечивающей компьютерные считывания дальности, высоты и пеленга на цель, как правило, каждые 10 с с радаром в режиме автоматического слежения. Все скорости полета были скорректированы с учетом влияния ветра путем вычитания вектора ветра на высоте полета птиц из вектора скорости полета птиц относительно земли. Ветер измерялся путем запуска и отслеживания заполненных водородом / гелием аэростатов с радарным отражателем.Средняя воздушная скорость, высота и вертикальная скорость полета рассчитывались для каждого трека, за исключением участков с запутанной траекторией полета. Высота была скорректирована по отношению к уровню моря путем добавления высоты антенны радара (10–185 м над уровнем моря в разных точках), а истинные воздушные скорости были уменьшены до эквивалентных воздушных скоростей ( U e ) относительно уровня моря. плотность воздуха, согласно стандартной атмосфере, изменение плотности воздуха с высотой [14,15].
Расчеты масштабирования и статистический анализ.
Редукционные регрессии по большой оси [16] для отношений масштабирования между U, , и и m, и Q, соответственно, были выполнены в Matlab с расчетами доверительных интервалов с помощью бутстрэппинга [33]. Расчеты сокращенных регрессий по большой оси на основе филогенетических независимых контрастов дополнительно описаны в Протоколе S2. Мы проверили возможное смещение, возникающее в результате включения видов с одним или несколькими следами, ограничив вычисления видами с минимум пятью или десятью следами.Результаты остались такими же, как показано на примере выборки из 56 видов с ≥10 следов в Таблице 1. Для 39 видов с ≥10 следов мы могли учесть внутривидовые вариации U e в соотношении к вертикальной скорости полета, компонентам встречного и бокового ветра и размеру стаи методом многомерной регрессии (статистически значимое влияние было обнаружено у 26 из этих 39 видов; неопубликованные данные). Ограничение анализа перехватом значений U e для этих 39 видов (с поправкой на нулевую вертикальную скорость, нулевой ветер и размер стаи, равный единице из уравнений множественной регрессии значимых переменных для каждого вида) по-прежнему дало такое же масштабирование результат (таблица 1).Общие линейные модели (рисунок 2) [34] были рассчитаны с U e в качестве зависимой переменной. Логарифмические значения использовались для U e , m, и Q . Филогенетическая группа и режим полета (ограниченный анализ этой предварительно оцененной переменной) рассматривались как фиксированные факторы. Сложные модели (различные комбинации или взаимодействия массы, соотношения сторон и филогенетической группы или нагрузки крыла, соотношения сторон и филогенетической группы) были представлены на рисунке 2 только в том случае, если AIC улучшился по сравнению с моделями с отдельными независимыми переменными [19].
Благодарности
Мы очень благодарны Инге Рудебек, которая участвовала во всех полевых радиолокационных исследованиях в Швеции и рассчитывала и компилировала результаты радиолокационного слежения, а также Бертилу Ларссону, который руководил работой радара, оборудованием и программным обеспечением для регистрации. Мы также благодарим нескольких дополнительных участников полевых работ: М. Грина, Г. А. Гудмундссона, А. Хеденстрёма и А. Ульфстранда. Радиолокационные услуги и реконструкции были выполнены Aerotech Telub. Ледокольные экспедиции в Арктику были организованы Секретариатом полярных исследований Швеции.Мы благодарим Б. Брудерера и А. Хеденстрема за комментарии к рукописи, а также Л. Ларссона и М. Ирестедта за обсуждения таксономической классификации птиц. Мы также благодарны Теунису Пирсме и анонимным рецензентам за ценные комментарии и предложения.
Вклад авторов
ТА и его лаборатория организовали и выполнили полевые радиолокационные работы. MR извлек и подготовил окончательный набор данных и выполнил анализ масштабирования. JB организовал информацию отслеживания в базу данных, включая информацию о биометрии.PGPE построила филогенетическое дерево и классифицировала виды на основные филогенетические группы, а ОН выполнила масштабный анализ на основе данных филогенетического контраста. TA, MR, JB, PGPE и OH участвовали в оценке и обсуждении результатов и написании рукописи.
Список литературы
- 1. Лайтхилл Дж. (1977) Введение в масштабирование передвижения животных. В: Pedley TJ, редактор. Масштабные эффекты передвижения животных. Нью-Йорк: Academic Press. С. 365–404.
- 2. Rayner JMV (1988) Форма и функции в птичьем полете. В: Джонстон РФ, редактор. Том 5, Современная орнитология. Нью-Йорк и Лондон: Plenum Press. С. 1–66.
- 3. Спеддинг Г.Р. (1992) Аэродинамика полета. В: Александр Р, редактор. Том 11, Успехи в сравнительной физиологии и физиологии окружающей среды. Берлин и Гейдельберг: Springer-Verlag. С. 52–111.
- 4. Pennycuick CJ (1969) Механика миграции птиц. Ibis 111: 525–556.
- 5. Greenewalt CH (1975) Полет птиц. Том 65, Trans Am Phil Soc New Series - часть 4. 67 с.
- 6. Теннекес Х. (1997) Простая наука о полете - от насекомых до гигантских самолетов. Кембридж (Массачусетс): MIT Press. 138 с.
- 7. Такер В.А. (1973) Метаболизм птиц во время полета: оценка теории. J Exp Biol 58: 689–709.
- 8. Rayner JMV (1979) Новый подход к механике полета животных. J Exp Biol 80: 17–54.
- 9. Rayner JMV (1995) Механика полета и ограничения на летные характеристики. Isr J Zool 41: 321–342.
- 10. Норберг У.М. (1990) Полёт позвоночных. Берлин: Springer-Verlag. 291 с.
- 11. Pennycuick CJ (1982) Полет буревестников и альбатросов (Procellariiformes), наблюдался в Южной Георгии и ее окрестностях. Фил Транс Р. Соц Лондон B 300: 75–106.
- 12. Эриксон PGP, Андерсон К.Л., Бриттон Т., Эльзановски А., Йоханссон США и др.(2006) Диверсификация новообразований: интеграция данных молекулярной последовательности и окаменелостей. Биол Лет 2: 543–547.
- 13. Хеденстрём А., Алерштам Т. (1995) Оптимальная скорость полета птиц. Фил Транс Р. Соц Лондон B 348: 471–487.
- 14. Pennycuick CJ (1989) Летные характеристики птицы: практическое руководство по расчетам. Оксфорд: Издательство Оксфордского университета. 153 с.
- 15. Брудерер Б., Болдт А. (2001) Летные характеристики птиц: I. Радиолокационные измерения скорости.Ibis 143: 178–204.
- 16. Харви PH, Пагель MD (1991) Сравнительный метод в эволюционной биологии. Оксфорд: Издательство Оксфордского университета. 239 с.
- 17. Джонсон Дж. Б., Омланд К. С. (2004) Выбор модели в экологии и эволюции. Тенденции Ecol Evol 19: 101–108.
- 18. Pennycuick CJ (1975) Механика полета. В: Фарнер Д.С., Кинг-младший, редакторы. Том 5, Биология птиц. Нью-Йорк: Academic Press. С. 1–75.
- 19. Hedenström A, Alerstam T (1992) Показатели лазания перелетных птиц как основа для оценки пределов топливной способности и работы мышц.Дж. Эксп. Биол. 164: 19–38.
- 20. Фогель С. (1994) Жизнь в движущихся жидкостях. Физическая биология потока. 2-е издание. Принстон: Издательство Принстонского университета. 467 с.
- 21. Pennycuick CJ (2001) Скорость и частота взмахов крыльев перелетных птиц по сравнению с расчетными контрольными показателями. J Exp Biol 204: 3283–3294.
- 22. Тобальске Б.В., Хедрик Т.Л., Диал К.П., Бивенер А.А. (2003) Кривые сравнительной мощности в полете птиц. Природа 421: 363–366.
- 23.Kvist A, Lindström Å, Green M, Piersma T, Visser GH (2001) Перевозка большого количества топлива во время продолжительного полета дешевле, чем ожидалось. Природа 413: 730–732.
- 24. Ward S, Möller U, Rayner JMV, Jackson DM, Bilo D, et al. (2001) Метаболическая мощность, механическая мощность и эффективность во время полета в аэродинамической трубе европейского скворца Sturnus vulgaris. J Exp Biol 204: 3311–3322.
- 25. Спеддинг Г.Р., Розен М., Хеденстрём А. (2003) Семейство вихревых следов, создаваемых дроздовым соловьем в свободном полете в аэродинамической трубе во всем естественном диапазоне скоростей полета.J Exp Biol 206: 2313–2344.
- 26. Розен М., Спеддинг Г.Р., Хеденстрём А. (2004) Взаимосвязь между кинематикой взмаха крыльев и структурой вихревого следа у соловья-дрозда. J Exp Biol 207: 4255–4268.
- 27. Hedenström A, Rosén M, Spedding GR (2006) Вихревые следы, создаваемые малиновкой Erithacus rubecula во время свободного полета в аэродинамической трубе. Интерфейс JR Soc 3: 263–276.
- 28. Тейлор Г.К., Наддс Р.Л., Томас А.Л.Р. (2003) Летающие и плавающие животные совершают круиз по числу Струхаля, настроенному на высокую энергоэффективность.Природа 425: 707–711.
- 29. Nudds RL, Taylor GK, Thomas ALR (2004) Настройка числа Струхаля для обеспечения высокой эффективности движения точно предсказывает, как частота взмахов крыльев и амплитуда гребков связаны и масштабируются с размером и скоростью полета у птиц. Proc R Soc Lond B 271: 2071–2076.
- 30. Спеддинг Г. Р., Хеденстрём А., Розен М. (2003) Количественные исследования следов свободно летающих птиц в аэродинамической трубе с низкой турбулентностью. Exp Fluids 34: 291–303.
- 31.Тобальске Б.В., Хедрик Т.Л., Бивенер А.А. (2003) Кинематика крыла полета птиц на скоростях. Дж. Авиан Биол 34: 177–184.
- 32. Алерстам Т., Гудмундссон Г.А. (1999) Модели миграции тундровых птиц: слежение за радиолокационными наблюдениями вдоль Северо-Восточного прохода. Арктика 52: 346–371.
- 33. Bohonak AJ (2004) RMA, программное обеспечение для сокращенной регрессии по главной оси, версия 1.17 [компьютерная программа]. Доступно: http://www.bio.sdsu.edu/pub/andy/rma.html. По состоянию на 1 июля 2006 г.
- 34. SPSS (2003) Версия 12.0.1 для Windows [компьютерная программа]. Доступно: http://www.spss.com. По состоянию на 1 ноября 2003 г.
Скорость и частота взмахов крыльев перелетных птиц по сравнению с расчетными ориентирами
Введение
Оптимальная скорость воздуха
Hedenström и Alerstam (Hedenström and Alerstam, 1995) утверждали, что разные оптимальные скорости воздуха могут быть определены для мигрирующей птицы в зависимости от того, является ли «валютой», которую птица пытается максимизировать, расстояние на единицу используемой энергии топлива, средняя скорость более или несколько этапов полета и промежуточных остановок, или различные другие возможности.С другой стороны, Алерстам и Хеденстрём (Alerstam and Hedenström, 1998) признали, что на практике сложно определить, пытается ли мигрирующая птица максимизировать одну валюту, а не другую, сравнивая наблюдаемую скорость с альтернативными теоретическими оптимальными скоростями. Не меньшая из трудностей заключается в том, что для оценки теоретических скоростей сначала необходимо рассчитать для птицы кривую мощности по сравнению со скоростью . Для этого требуются некоторые морфологические данные, которые никогда не доступны для каждой отдельной птицы, наблюдаемой в поле.Можно использовать видовые средства, но в случае радиолокационных наблюдений даже вид часто вызывает сомнения. Даже когда доступны точные данные для измерения массы тела и крыльев, как в случае птицы, летящей в аэродинамической трубе, точная форма кривой мощности на высоких скоростях неизвестна. Признавая, что это критически важно для оценки оптимальных скоростей, Алерстам (Alerstam, 2000) предложил способ изменения кривой мощности для покрытия более высоких скоростей, но необходимы дополнительные экспериментальные данные, прежде чем пересмотренная модель может быть использована с уверенностью.
Расчетные ориентиры
Целью настоящего проекта является установление физических «эталонов», по которым можно будет измерять продуктивность диких птиц, без каких-либо гипотез относительно ожидаемых значений, при которых птицы «должны» летать. Основой для расчета контрольных показателей, включая сделанные предположения и принятые в настоящее время значения переменных, является базовая программа Flight.bas, опубликованная в Интернете (http://detritus.inhs.uiuc.edu / wes / pennycuick.html) и работает под управлением QBasic. Flight.bas основан на механике полета (а не статистике) и реализует физическую модель, описанную Pennycuick (Pennycuick, 1975; Pennycuick, 1989), которая сама является заимствованной из классической аэронавтики. Если экспериментальные результаты не согласуются с прогнозами программы, Flight.bas позволяет отследить источник несоответствия, чтобы исправить значения, принятые для переменных, которые нельзя измерить напрямую. Например, Pennycuick et al. (Pennycuick et al., 1996b) показали, что минимальная силовая скорость ( V, mp ) может быть получена эмпирически из измерений частоты взмахов крыльев у птиц, летящих в аэродинамической трубе с низкой турбулентностью, и они использовали это как проверку значения V. mp предсказано Flight.bas. Измеренные и рассчитанные значения V mp совпали у двух очень разных птиц, дрозда-соловья ( Luscinia luscinia , Turdidae) и чирка ( Anas crecca , Anatidae), когда коэффициент сопротивления тела C Db был установлен на 0.08, но программа занижала оценку V mp , когда для C Db использовались ранние, намного более высокие оценки. Эти аномально высокие значения, в диапазоне от 0,25 до 0,40, были получены путем измерения сопротивления замороженных бескрылых тел птиц и, как уже было известно, связаны с массивным отрывом потока, который, по-видимому, не наблюдается у живых птиц. Это наблюдение увеличивает уверенность, с которой вычисленные значения V mp могут использоваться для сравнения со скоростями, наблюдаемыми в полевых условиях.Использование слишком большого значения для C Db приводит к недооценке минимальной скорости и создает иллюзию, что дикие птицы летают с более высоким кратным V mp , чем они на самом деле.
Избранные ориентиры для воздушной скорости и частоты взмахов крыльев
V mp была выбрана в качестве эталонной скорости воздуха не потому, что ожидается, что любая птица будет летать с этой скоростью во время миграции, а потому что это единственная скорость на кривой мощности, для которой расчетные значения имеют некоторое экспериментальное подтверждение (см. Выше ).Кроме того, Лайтхилл (Lighthill, 1977) указал, что V mp является нижней границей диапазона скоростей, доступного для крейсерского полета с двигателем, и что длительный полет с двигателем на более низких скоростях практически невозможен. Причины этого являются механическими (а не физиологическими) и были дополнительно разработаны Pennycuick (Pennycuick, 1997). V mp был рассчитан для каждого вида по следующей формуле, которая адаптирована из Pennycuick (Pennycuick, 1975) и используется в Flight.bas:
1
, где м, - масса тела, г, - ускорение свободного падения, b - размах крыла, S b - лобовая площадь корпуса, C Db - коэффициент лобового сопротивления кузова, а k - коэффициент наведенной мощности. Установка плотности воздуха на фиксированное значение ρ SL , соответствующее уровню моря в теоретической стандартной атмосфере, приводит к расчетному «эквивалентному» значению для V mp .«Истинные» воздушные скорости, наблюдаемые в полевых условиях, должны быть «уменьшены до уровня моря» (см. Ниже), прежде чем их можно будет сравнить со значением, вычисленным по уравнению 1.
Эталонная частота биения крыльев ( f ref ) была рассчитана по формуле:
2
, где S - площадь крыла, определенная Пенникуиком (Pennycuick, 1999). Как и значение V mp , вычисленное по уравнению 1, f ref является эквивалентным значением уровня моря, поскольку оно основано на плотности воздуха на уровне моря (ρ SL ).Как и уравнение 1, эта формула содержит только физические переменные. Он был получен Пенникуиком (Pennycuick, 1990) из соображений размерности и модифицирован Пенникуиком (Pennycuick, 1996a) в свете полевых наблюдений за частотами взмахов крыльев.
Требования к полевым данным
Чтобы получить полевые данные для сравнения с эталонными показателями, требуется поток мигрирующих птиц, каждая из которых может быть идентифицирована до вида, для которого доступны измерения массы тела, размаха и площади крыла.Это требование ограничивает наблюдение местами и сезонами, где топография вызывает концентрацию миграционного потока и где мигранты летают достаточно низко для визуальной идентификации. Это, к сожалению, теряет преимущество наблюдений с помощью радара слежения, когда можно наблюдать высоко летающих птиц, которые определенно стремятся к полету на большие расстояния. Наблюдая только за полетом на малых высотах, трудно быть уверенным, какие птицы направляются к какому-либо отдаленному пункту назначения, а какие ищут корм на месте или останавливаются в районе исследования по маршруту в другое место.Выбирая район, который, как известно, является точкой концентрации для миграции, можно надеяться, что по крайней мере некоторые из наблюдаемых птиц мигрируют, чего не было в более ранних проектах с использованием аналогичных методов (Pennycuick, 1990; Pennycuick, 1996a). Помимо возможности визуальной идентификации каждой птицы, оптические методы наблюдения с близкого расстояния имеют важное преимущество, заключающееся в том, что ветер можно измерять сразу после каждого измерения скорости. Это обеспечивает гораздо лучшее временное разрешение, чем это возможно при отслеживании подъемов воздушного шара с интервалами в несколько часов, что обычно является единственным вариантом при радиолокационных наблюдениях.Кроме того, ветер можно измерить в точке, близкой к тому, где летит птица.
Материалы и методы
Место исследования и период
Птицы наблюдались с 3 по 15 октября 2000 г. с обзорной площадки напротив маяка Фальстербо, на краю дюн (55 ° 23,015 'северной широты, 12 ° 48,917' восточной долготы). Полуостров Фальстербо, расположенный на юго-западе Швеции, является местом концентрации мигрантов многих видов, покидающих Швецию осенью. Карлссон (Karlsson, 1992) описал этот район и исследования, проводившиеся там с момента основания в 1955 году станции кольцевания птиц на маяке, а также суммировал количество и время миграции различных видов птиц через этот район.Измерения массы тела, размаха крыльев и площади крыла приведены для 16 исследуемых видов в таблице 1. Эти измерения были взяты из базы данных, обобщенной Pennycuick (Pennycuick, 1999), или из более поздних добавлений к этой базе данных, за исключением данных для Columba palumbus. пришлось взять из Greenewalt (Greenewalt, 1962), потому что исходных измерений не было.
Измерение путевой скорости
Для измерения путевой скорости пролетающих птиц использовались два метода.
(i) Leica Vector - это пара биноклей 7 × 42 со встроенным лазерным дальномером, а также магнитным датчиком, который определяет направление, в котором направлен бинокль, и датчиком силы тяжести, определяющим угол места. Встроенные магниты-компенсаторы для датчика азимута могут быть отрегулированы автоматической системой при настройке прибора для устранения местных магнитных возмущений. При использовании инструмент наводится на цель с помощью перекрестия в центре поля и нажимается кнопка.Дальномер измеряет расстояние и отправляет результат, за которым следуют показания датчиков азимута и возвышения, через , последовательный порт вывода. Сообщение об ошибке отправляется, если дальномер не может обнаружить возвращающийся световой импульс. Вектор хорошо работал с крупными птицами светлого окраса, уверенно летящими на расстоянии 100–600 м от наблюдателя, но редко мог производить измерения дальности на мелких птицах.
Номинальная точность показания диапазона, выдаваемого вектором, составляла ± 0.5 мес. Повторное определение расстояния до статических целей привело к стандартным отклонениям от нуля на 398 м (20 измерений, все равно) до ± 0,37 м на 149 м. Такое же стандартное отклонение (± 0,24 м) было измерено как на 92 м, так и на 779 м. Таким образом, повторяемость измерения дальности была лучше, чем размах крыльев большинства исследованных видов (таблица 1). Номинальная точность двух угловых энкодеров составляла 10 –4 рад (0,0057 °). Стандартные отклонения составляли от 0,10 до 0,14 ° по азимуту и от 0.От 050 до 0,073 ° по углу места. Эти значения являются реалистичным показателем точности используемого вектора, поскольку считывание диапазона невозможно было получить, если прибор не был точно выровнен по птице. Угловая погрешность ± 0,2 ° на дальности 500 м будет соответствовать ошибке позиционирования ± 1,7 м, но повторяемость на статических целях была лучше, чем эта. Были предприняты попытки вызвать ошибки в датчике силы тяжести путем вращения инструмента с резкими пусками и остановками, но ошибок не было обнаружено.Чтобы свести к минимуму такие эффекты, инструмент был установлен на тяжелом фотографическом штативе Gitzo с наклонно-поворотной головкой с гидравлическим демпфированием, предназначенной для использования с видеокамерами и обеспечивающей очень плавное вращение.
(ii) Орнитодолит измеряет расстояние с помощью дальномера оптического совпадения с базой 25 см, а азимут и углы возвышения с помощью восьмиразрядных оптических кодеров. Этот прибор, его калибровка и точность были описаны Pennycuick (Pennycuick, 1982a). Для данного проекта был построен новый интерфейс на основе микроконтроллера PIC 16C74, который считывает показания датчиков при нажатии кнопки и выводит показания в последовательной форме.Орнитодолит использовался для отслеживания мелких птиц на расстоянии 50–120 м от наблюдателя.
Какой бы прибор ни использовался, данные отправлялись в последовательный порт портативного компьютера (MBC Proteus) и считывались вместе со временем с часов реального времени компьютера программой, написанной на Microsoft QBasic. «Тиковый» период часов составил 54,925 мс, что является улучшением по сравнению с более ранним интерфейсом орнитодолита, разрешение по времени которого составляло 100 мс. Данные были преобразованы из полярных координат в декартовы (Pennycuick, 1982b) и записаны как трехмерные точки в пространстве с временной привязкой.Как и в предыдущих проектах орнитодолита, «пробег» определялся как серия из двух или более таких точек, а «наблюдение» представляло собой измерение путевой скорости (горизонтальной и вертикальной), полученное путем сравнения каждой точки с предыдущей. Таким образом, количество наблюдений в каждом прогоне было на единицу меньше, чем количество точек. Количество прогонов и наблюдений для каждого из 16 исследуемых видов показано в Таблице 1. Данные были сохранены в полевых условиях на жестком диске и были в одном и том же формате, независимо от того, источником данных был вектор или орнитодолит.
Измерение ветра
Аппаратное обеспечение анемометра состояло из узла вихревой чашки и лопасти от Maplin Electronics, снабженного дисками оптического кодировщика, самодельными из литовой пленки. Направление ветра измерялось семибитным угловым энкодером с кодировкой Грея с шагом 5 °. Скорость ветра измерялась с помощью 36-сегментного непрерывно вращающегося диска путем подсчета секторов, проходящих через оптический датчик за фиксированный период. Датчик скорости ветра был откалиброван в аэродинамической трубе Лунда (Pennycuick et al., 1997), чтобы получить истинную (не эквивалентную) скорость воздуха в мс –1 . Оба датчика считывались микроконтроллером PIC 16C71, который отправлял показания с интервалом 2,3 с. через UHF-радиоканал на приемник, который был подключен ко второму последовательному порту на компьютере (Socket S-I / O). Этот порт активировался управляющей программой всякий раз, когда цикл был завершен и сохранен. Скорость и направление ветра были автоматически получены и записаны как часть данных для пробега.
Блок анемометра был установлен на мачте на высоте 3 м над землей.Радиосвязь устраняла необходимость в кабеле анемометра, так что мачту можно было расположить вдали от любых препятствий с наветренной стороны. Помимо сада маяка, который поддерживал рощу деревьев, в непосредственной близости находилось ровное и относительно свободное поле для гольфа с восточной и южной сторон и воды Эресунда с запада и севера. В любом направлении не было холмов, достаточно высоких, чтобы вызвать ветровые волнения. Поскольку высота полета птиц находилась в пределах 200 м от поверхности, корректировка направления ветра не требовалась, но скорость ветра корректировалась в соответствии с обычным уравнением градиента ветра:
3
, где V h3 - скорость ветра на высоте полета птицы ( h 2 ), V h2 - измеренная скорость ветра на анемометре, высота которого составляет h 1 , а h r - это «высота шероховатости», которой можно присвоить значение от 10 –4 м для гладкой стекловидной поверхности до 10 –2 м для очень шероховатой поверхности (Sutton, 1953).«Среднее» значение 10 –3 м использовалось на всем протяжении h r . Анемометр с вращающейся чашей измеряет истинную (не эквивалентную) скорость ветра, которая после поправки на высоту полета была векторно вычтена из измеренной скорости относительно земли, чтобы дать оценку истинной скорости воздуха. Скорости ветра с поправкой на высоту полета варьировались от 0 до 14,3 м с –1 .
Частота колебаний крыла
Аналоговое видео было записано в полевых условиях с помощью видеокамеры Panasonic NV-S7B на кассетах S-VHS-C в европейском формате PAL, в котором 50 «полей», каждое с половиной горизонтальных строк полного «кадра». ', записываются каждую секунду с равными интервалами времени 20 мс.Частоты биений крыльев измерялись с копии оригинальной ленты, на которой были пронумерованы отдельные поля (Pennycuick, 1996a). Каждая последовательность взмахов крыльев воспроизводилась в замедленном или пошаговом режиме, по одному полю за раз, начиная и заканчивая последовательность в одной и той же точке цикла взмахов крыльев. Время для последовательности, а следовательно, и частота, определялись по номерам первого и последнего полей. Стандартное отклонение частоты взмахов крыльев оценивалось путем определения среднего и стандартного отклонения его обратной величины, периода взмахов крыльев.Причина этого и метод расчета были объяснены Пенникуиком (Pennycuick, 1990).
В случае устойчивого махающего полета часто можно было измерить несколько последовательностей махов от конца до конца, причем каждая последовательность начинается в том поле, где закончилась предыдущая последовательность. Как и в предыдущих исследованиях, предпочтительной точкой в цикле для начала и окончания каждой последовательности был момент в начале хода вниз, когда крылья испытывают нагрузку. Этот переход резкий и его можно распознать на взглядах с разных сторон.Однако при парящем и ограничивающем полете некоторые птицы имели тенденцию начинать последовательность с частичного движения вниз от планирующего положения, и было обнаружено, что более практичным было использовать положение «полностью опущено» для начала и завершения последовательности взмахов. Из-за этого время от первой до последней точки полного опускания последовательности было короче на один период взмахов крыльев, чем истинная длительность последовательности, и это нужно было учитывать при измерении «доли мощности» при планировании с закрылками. или ограничивающий полет, i.е. доля времени, затраченного на хлопанье. «Частота взмахов крыльев» в этих прерывистых стилях полета относится к частоте в последовательности непрерывных взмахов, а не к среднему значению за периоды взмахов и без них. Количество полученных записей и подсчитанных взмахов крыльев показано для каждого из 16 исследуемых видов в таблице 1.
Редукция данных
Ожидается, что как воздушная скорость, так и частота взмахов крыльев будут функциями плотности воздуха, измеренное значение которой ρ менялось от одного полета к другому.Плотность воздуха в месте нахождения наблюдателя сначала определялась по температуре окружающего воздуха и атмосферному давлению, а затем корректировалась с учетом измеренной разницы в высоте между наблюдателем и птицей. Формулы для этих операций были даны Pennycuick (Pennycuick, 1999). Плотности воздуха на уровне моря в стандартной атмосфере было присвоено фиксированное значение (ρ SL ) 1,23 кг м –3 . σ определяется как отношение плотности окружающего воздуха к стандартной:
4
В соответствии с авиационной конвенцией, наблюдения за воздушной скоростью и частотой взмахов крыльев были «уменьшены» до значений, которые преобладали бы на уровне моря, согласно теоретическим ожиданиям, до того, как были предприняты попытки сравнения.Поскольку ожидается, что скорость воздуха будет изменяться обратно пропорционально квадратному корню из плотности воздуха (уравнение 1), измеренная «истинная» скорость воздуха ( V t ) была уменьшена до «эквивалентной» скорости воздуха на уровне моря ( V e ) таким образом:
5
Эквивалентная частота биений крыльев ( f e ) была получена аналогичным образом путем уменьшения измеренной истинной частоты биений крыльев ( f t ) до значения на уровне моря.Поскольку ожидается, что частота взмахов крыльев будет меняться обратно пропорционально степени плотности воздуха 3/8 (уравнение 2), уравнение уменьшения в этом случае будет:
6
Эффект от приведения данных к уровню моря был небольшим, поскольку плотность воздуха на высоте полета изменялась только от 1,19 до 1,26 кг · м –3 , давая значения σ от 0,98–1,02. Эти сокращения будут иметь больший эффект в ситуации, когда птицы наблюдаются в более широком диапазоне высот, как в исследованиях с помощью радиолокатора слежения.В этом случае (а также, в принципе, в данном проекте) нередуцированные наблюдения не сопоставимы ни с эталонным значением, рассчитанным для фиксированной высоты, ни друг с другом.
Результаты
Расчетные и наблюдаемые значения
В таблице 2 показаны средние наблюдаемые эквивалентные воздушные скорости и частоты взмахов крыльев для каждого из 16 исследуемых видов вместе с оценками минимальной скорости движения ( V mp ) для каждого вида, рассчитанными по уравнению 1, и эталонной частотой взмахов крыльев ( f ref ) из уравнения 2.Значения морфологических переменных для каждого вида приведены в таблице 1, а значения, используемые для других переменных в уравнениях 1 и 2, и их предполагаемые неопределенности приведены в таблице 3.
На рисунках 1 и 2 наблюдаемые средние значения воздушной скорости и частоты взмахов крыльев сравниваются с эталонными значениями, рассчитанными для каждого вида из уравнений 1 и 2, путем нанесения отношения наблюдаемого значения к расчетному на горизонтальной плоскости. шкала. Наблюдаемые значения являются средними значениями для образцов и сопровождаются в таблице 2 стандартными отклонениями, но эталонные значения были получены непосредственно из измерений в таблицах 1 и 3.Эти оценки не имеют стандартных отклонений как таковых, поскольку они не являются выборочной статистикой, но каждая имеет «неопределенность». Это происходит из-за неопределенностей значений, используемых для отдельных переменных в правой части уравнений 1 и 2, каждая из которых вносит свой вклад в неопределенность результата в левой части. Неопределенности V mp и f ref были рассчитаны по методу Спеддинга и Пенникуика (Spedding and Pennycuick, 2001) и перечислены в таблице 2.Процедура является обычной в физических науках, но основана на принципе, отличном от принципа вычисления стандартного отклонения выборки. Было бы неправильно использовать эти оценки неопределенности в статистических тестах значимости, поскольку они не удовлетворяют допущениям, лежащим в основе тестов этого типа. В любом случае не существует исходной гипотезы относительно скоростей, с которой «должны» летать птицы, которую можно было бы отклонить путем проверки значимости.
Рис. 1.Наблюдаемая эквивалентная скорость воздуха ( V obs ) с горизонтальными полосами стандартного отклонения, нормализованная путем деления на рассчитанную минимальную скорость вращения ( V mp ).Квадраты предназначены для видов, которые летали на крыльях, кресты для взмахов и круги для прыжков. Данные из таблицы 2.
Рис. 2.Как на рис. 1, но показывает отношение наблюдаемой эквивалентной частоты биений крыла к расчетной ( f obs / f ref ). Данные из таблицы 2.
Стиль полета
Можно выделить три различных стиля полета: непрерывный взмахи крыльями (для большинства видов), скольжение с закрылками и прыжок.Три вида хищных птиц в выборке неизменно летали путем взмахов крыльев, т. Е. Поочередно взмахивая крыльями и планируя на короткие периоды времени. Зяблик ( Fringilla coelebs ) неизменно летел, прыгая, то есть поочередно взмахивая крыльями, затем свободно падая с закрытыми крыльями, в то время как скворец ( Sturnus vulgaris ) иногда летел, прыгая, а иногда и продолжая хлопать крыльями. Скворцов не было замечено ни визуально, ни на видео.
Эквивалентная скорость воздуха
Виды расположены на рис.1 в порядке убывания их «относительной воздушной скорости», что означает отношение наблюдаемой средней эквивалентной воздушной скорости к расчетной минимальной силовой скорости, обе из таблицы 2. Относительная воздушная скорость, равная 1 (сплошная вертикальная линия), означает, что наблюдаемая средняя скорость равнялась расчетному значению V mp . Неопределенность V mp , также приведенная в Таблице 2, варьировалась у разных видов животных в пределах от 8 до 12% от расчетного значения, что приблизительно обозначено вертикальными пунктирными линиями в точке 0.90 и 1.10. На рис. 1 использованы разные символы для классификации стилей полета, используемых каждым видом. Крестиками обозначены виды, которые летели непрерывным машущим полетом, и они доминируют в средней части рис. 1 с наблюдаемыми относительными скоростями воздуха от 0,83 до 1,27. Три вида, летевшие с помощью закрылков (квадраты), сгруппированы в нижней части таблицы с относительной скоростью воздуха до 0,73, в то время как два вида, которые летели ограничиваясь (кружки), находятся вверху, со средней скоростью воздуха. почти вдвое V mp .
Эффект ветра
«Попутный ветер» обычно определяется как скалярная разница между путевой скоростью и истинной воздушной скоростью. Величина этой разницы отражает степень, в которой ветер помогает или мешает птице, независимо от того, совпадает ли ветер с курсом птицы или под углом к нему. «Эффект ветра» означает, что птица, скорость которой относительно земли меньше ее воздушной скорости, обычно реагирует увеличением воздушной скорости, что приводит к отрицательной корреляции между воздушной скоростью и таким определенным «попутным ветром» (Pennycuick, 1982b).На рис. 3 показаны такие графики для трех видов с разными стилями полета. Зяблик показал очень сильное ветровое воздействие со скоростью воздуха от 8,64 м с –1 (1,1 V mp ) до 24,3 м с –1 (3,1 V mp ), что было самая высокая скорость, измеренная у любого вида в этом исследовании, абсолютная или относительно V mp . Высокая средняя скорость зяблика (1,96 V mp ) отражает его широкий диапазон скоростей и его способность увеличивать скорость при полете против встречного ветра.Соответствующая линия на фиг. 3A пересекает нулевой ветер (вертикальная пунктирная линия) при скорости воздуха только чуть выше V mp . У обыкновенного канюка ( Buteo buteo ), который летел с помощью закрылков, почти все измерения скорости были ниже расчетной минимальной скорости, тогда как у баклана ( Phalacrocorax carb ), который летал, взмахивая крыльями, большинство было ниже V мп но некоторые были и выше. Коэффициент корреляции был отрицательным и высокодостоверным ( P ≪0.01) у каждого из этих трех видов, хотя некоторые другие виды, у которых количество образцов было небольшим, не показали значимой корреляции.
Рис. 3.Эффект ветра у видов, которые летали путем прыжков (A), непрерывных хлопков (B) и скольжения с закрылками (C) с уменьшенными линиями больших осей. «Хвостовой ветер» - это скалярная разница между путевой и воздушной скоростью. Горизонтальные пунктирные линии показывают минимальную скорость вращения ( V mp ) и минимальную скорость планирования ( V min ) при максимальном коэффициенте подъемной силы, равном 1.6.
Частота колебаний крыла
На рис. 2 виды расположены в порядке убывания «относительной частоты взмахов крыльев» для каждого вида, т.е. отношения наблюдаемой эквивалентной частоты взмахов крыльев к расчетному значению f ref из таблицы 2. Поскольку на рис. 1 два ограничивающих вида (кружки) занимают два верхних места. Зяблик был единственным видом, у которого наблюдаемая частота взмахов крыльев была намного выше, чем расчетное значение f ref , а стандартное отклонение также было намного выше, чем у любого другого вида.Три вида закрылков (квадраты) не сгруппированы в какой-либо определенной части таблицы.
Обсуждение
Птицы, летящие медленнее
V mpХотя не ожидалось, что какая-либо птица будет летать со скоростью, меньшей, чем значение V mp , рассчитанное для этого вида, наблюдаемые скорости были равномерно разделены ниже и выше V mp . Восемь из 16 видов летали со средней скоростью от 0 до 0.73 и 0,99 V mp , а остальные восемь видов между 1,02 и 1,96 V mp . Средние скорости трех видов были более чем на одно стандартное отклонение ниже V mp . Пилоты называют это «полетом на обратной стороне кривой мощности», а непрерывный полет на таких низких скоростях затруднен как для птиц, так и для самолетов (Pennycuick, 1997). Однако три из четырех видов с наименьшими относительными скоростями воздуха были хищниками, которые летали с помощью закрылков.В их случае средняя скорость ниже V mp объяснима при условии, что птица способна скользить медленнее, чем V mp . Расчетные минимальные скорости планирования трех видов планеристов, перепелятников ( Accipiter nisus ), канюка ( Buteo buteo ) и красного коршуна ( Milvus milvus ), исходя из максимального коэффициента подъемной силы 1,6 , выраженные как кратные V mp , находились в пределах 0.49 и 0,57. Наблюдаемые средние относительные скорости в течение нескольких циклов скольжения закрылков составляли от 0,73 до 0,91. Вероятное объяснение явной аномалии средних скоростей ниже V mp состоит в том, что птица замедляется во время фазы планирования значительно ниже V mp , но начинает махать, прежде чем приблизится к минимальной скорости (сваливанию), затем разгоняется при махах до V mp или чуть выше. Планирование закрылков включает цикл изменения скорости с максимальной скоростью около V mp и средней скоростью ниже V mp .Термин «волнообразный» полет для планирования с закрылками (Rayner, 1977; Rayner, 1985) вводит в заблуждение, и его следует избегать. Скорость колеблется, но высота обычно не меняется, по крайней мере, недостаточно, чтобы ее можно было легко различить.
Низкая относительная скорость (0,82), наблюдаемая у лебедя-шипуна ( Cygnus olor ) при взмахе крыльев, является аномальной и не может быть объяснена таким образом. Скорости были очень постоянными (низкое стандартное отклонение), вероятно, потому, что одна и та же небольшая группа лебедей летала взад и вперед вдоль берега.Несоответствие в этом направлении могло бы произойти, если бы значение, принятое для коэффициента наведенного сопротивления ( k = 1,2), было бы слишком высоким, но эффект невелик. Наименьшее возможное значение - k = 1.0 для невероятного приводного диска без потерь. Это снизит оценку V mp с 19,4 до 18,5 м с –1 и увеличит относительную скорость с 0,82 до 0,86, оставив аномалию неразрешенной. Этим лебедям требуется длительный разбег и некоторое время и некоторое расстояние, чтобы разогнаться до постоянной скорости полета, и иногда они приземляются на воду возле маяка в пруду за пляжем.Намерение состояло в том, чтобы измерить их скорость только тогда, когда они стабильно летят, но возможно, что средняя скорость была непреднамеренно смещена вниз, сделав некоторые наблюдения до того, как лебеди разогнались до V mp после взлета или когда они начали. сбавить скорость для возможной посадки.
Птицы, летящие со скоростью около
V или чуть выше mpНад ястребом-перепелятником ( Accipiter nisus ) на рис.1 представлены шесть видов, средние эквивалентные скорости воздуха которых находились в пределах одного стандартного отклонения от расчетных V mp , затем четыре вида с относительной скоростью воздуха 1,20–1,27. Все эти виды летали непрерывным взмахом крыльев. У птиц, летящих немного быстрее, чем V mp , нет физических аномалий, но различные теоретические оптимальные скорости, предложенные Хеденстремом и Алерштамом (Hedenström and Alerstam, 1995), были бы намного выше. Самая низкая из них - максимальная скорость ( V mr ), при которой эффективное отношение подъемной силы к лобовому сопротивлению проходит через максимум.Оценки отношения V mr к V mp для видов в выборке, рассчитанные Flight.bas, варьируются от 1,60 до 1,89, но следует отметить, что оценки V mr менее надежны, чем V mp , потому что значение V mr зависит от точной формы кривой мощности, которая малоизвестна. Недавние физиологические данные показывают, что кривая химической мощности имеет более U-образную форму, чем Flight.базовый прогноз (Квист и др., 1998). V MR представляет собой широкий пик эффективного отношения подъемной силы к лобовому сопротивлению в области кривой мощности, где сама мощность резко возрастает, и прямые затраты с точки зрения топливной энергии ни в коем случае не являются единственным фактором, влияющим на выбор крейсерской скорости (Pennycuick, 1997). Результаты показывают, что виды, летящие в устойчивом колеблющемся полете, летели со скоростью выше V mp , но намного ниже V mr , что во многом аналогично тому, что наблюдалось в предыдущих проектах (Pennycuick, 1990; Pennycuick, 1996a), при этом считалось, что птицы совершают короткие локальные перелеты, а не мигрируют.Возможно, что те же самые птицы будут летать с более высокими эквивалентными воздушными скоростями, если будут совершать миграцию на больших высотах, но для этого истинные воздушные скорости должны быть еще выше (уравнение 5).
Птицы, которые летели намного быстрее, чем
V mp : ограничиваяНаконец, два ограничивающихся вида в верхней части диаграммы, скворец ( Sturnus vulgaris ) и зяблик ( Fringilla coelebs ), летели со средней относительной скоростью воздуха 1.70 и 1,96 соответственно, более чем на два стандартных отклонения выше V mp в каждом случае. Эти скорости близки (несколько неопределенные) к оценкам V mr из Flight.bas для этих двух видов. Хеденстрём и Алерстам (Hedenström and Alerstam, 1992) отметили, что мигрирующие зяблики и некоторые другие мелкие виды на самом деле летели быстрее, чем их оценка для V mr , но в то время они использовали чрезмерно высокие значения коэффициента сопротивления тела и, таким образом, занижая V mp и V mr (см. выше).На рис. 3А показано, что высокая средняя скорость зяблика является результатом его способности увеличивать скорость при полете против встречного ветра, но рис. 1 предполагает, что два прячущихся вида были единственными, кто мог это сделать.
Механические характеристики ограничивающего полета были определены Лайтхиллом (Lighthill, 1977) и в дальнейшем развиты Рейнером (Rayner, 1977; Rayner, 1985). Ограничивающий полет требует циклического изменения восходящего ускорения, в отличие от горизонтального полета или полета с закрылками, которые этого не делают.На рис. 4 показан упрощенный связанный цикл, состоящий из двух регулярно повторяющихся фаз без чистого изменения высоты. В «баллистической фазе» птица держит свои крылья сложенными относительно своего тела, делая обтекаемую веретенообразную форму, которая, как предполагается, не создает подъемной силы. Птица находится в свободном падении, и траектория полета изгибается вниз. Оставшаяся часть цикла - это «фаза мощности», во время которой птица машет крыльями, а траектория полета изгибается вверх. Увеличение лобовой площади из-за обертывания крыльев вокруг тела кажется небольшим, и предполагается, что сопротивление в баллистической фазе такое же, как у тела без крыльев.
Рис. 4.Поскольку она падает в свободном падении во время баллистической фазы, прыгающая птица должна подтягиваться во время фазы взмахов, эффективно увеличивая силу тяжести ( g ) с коэффициентом 1/ q , где q - доля мощности (доля времени, потраченного на хлопанье).
В непрерывном махающем полете механическая мощность ( P mech ) представлена в Flight.bas как сумма трех составляющих: наведенная мощность ( P ind ), мощность профиля ( P pro ) и паразитная сила ( P par ), из которых первые два являются функциями силы тяжести, а третье - нет:
7
При ограничении все три компонента требуются во время фазы питания, но только паразитная мощность требуется во время фазы баллистики.Сила паразита - это скорость, с которой работа рассеивается на преодоление аэродинамического сопротивления тела. Он не включает работу, проделанную против компонента веса в соответствии с волнообразной траекторией полета, которая сводится к нулю в течение полного цикла, как в американских горках.
Обозначая «долю мощности» или долю от общего времени цикла, затраченного на взмахи, как q , ускорение вверх во время фазы мощности должно быть таким, чтобы вес тела увеличивался на «коэффициент нагрузки», равный 1 / q (Лайтхилл, 1977).Фактически величина силы тяжести, с которой птица должна поддерживать свой вес во время фазы мощности, составляет не г , а г ′ , где:
8
Индуцированная и профильная составляющие мощности, которые являются функциями g в горизонтальном полете, становятся функциями g ′ в ограничении, но требуются только для части времени q . Сила паразита ( P, пар. ) не зависит от силы тяжести и требуется все время, независимо от того, машет птица или нет.Таким образом, средняя механическая мощность за весь связанный цикл составляет:
.9
На рис. 5 показано влияние изменения доли мощности на кривые механической мощности и эффективного отношения подъемной силы к сопротивлению, рассчитанного с помощью Flight.bas для зяблика на основе данных в Таблице 1 и Таблице 3. Кривые иллюстрируют те же общие выводы, что и были достигнуты Райнером (Rayner, 1985), т. е. мощность, необходимая для полета с любой заданной скоростью, увеличивается с уменьшением доли мощности, в то время как эффективное отношение подъемной силы к лобовому сопротивлению уменьшается, а также что и V mp и V MR переключайтесь на более высокие скорости по мере уменьшения доли мощности.Как отметил Рейнер (Rayner, 1985), ограничение не дает никаких прямых преимуществ с точки зрения требуемой мощности или расстояния полета на единицу потребляемой энергии.
Рис. 5.Расчетные кривые для зяблика Fringilla coelebs : (A) механической мощности и (B) эффективного отношения подъемной силы к сопротивлению (на основе метаболической мощности) для различных значений доли мощности ( q ). Эти кривые были расширены до 25 м с –1 , поскольку зяблики наблюдались, летящие почти с этой скоростью, но расчет на таких высоких скоростях ненадежен.
Частота колебаний крыла
Как отмечалось выше, уравнение 2 для эталонной частоты биений крыла f ref основано на физических соображениях и не включает никаких коэффициентов регрессии или даже числовых коэффициентов преобразования. Тем не менее, средняя наблюдаемая частота взмахов крыльев, при делении на f ref , дает относительные частоты взмахов крыльев от 0,81 до 1,05 у 14 из 16 видов. f ref был эталоном, рассчитанным на основе размерного аргумента.Хотя не обязательно было предсказывать частоту взмахов крыльев, рис. 2 показывает, что на самом деле f ref действительно предсказал частоту взмахов крыльев и взмахов закрылков, а также что два ограничивающих вида имели самые высокие частоты взмахов крыльев относительно f исх. . Оба ограничивающих вида также показали большие стандартные отклонения, чем любые другие 14 видов. Наблюдаемая средняя эквивалентная частота находилась в пределах одного стандартного отклонения от f ref в случае скворца, но более четырех стандартных отклонений выше у зяблика.
Контрольные значения f ref были рассчитаны из уравнения 2 с использованием фиксированного значения g в Таблице 3. Однако частота взмаха крыльев была определена выше как частота во время фазы взмахов прыжка (а не среднее значение). частота в связанном цикле), и было также отмечено, что, когда птица подтягивается в фазе взмахов, значение силы тяжести увеличивается на коэффициент нагрузки 1/ q , где q - доля мощности.Это можно выразить, изменив уравнение 2 ограничивающего (но не планирующего) полета на:
10
с использованием г / q вместо г для силы тяжести. Это сильный эффект. Средний коэффициент мощности зябликов составил 0,35, что составляет коэффициент нагрузки 2,9, т.е. зяблики «тянули 2,9 г », как сказали бы пилоты. Это увеличивает частоту биений крыла, согласно уравнению 10, примерно в √2 раз.9 или 1,7. Доли мощности у скворца варьировались от 0,57 до 1,0, давая коэффициент нагрузки от 1,8 до 1,0 и увеличивая частоту взмахов крыльев в 1,3 раза.
На рис. 6 показаны наблюдаемые частоты взмахов крыльев в зависимости от доли мощности как для скворца, так и для зяблика. Кривые показывают частоту взмахов крыльев, предсказанную для каждого вида, как функцию доли мощности из уравнения 10, в то время как горизонтальные линии представляют собой предсказания из уравнения 2, представляя гипотезу об отсутствии эффекта от увеличения силы тяжести при ограничении.Отношение среднеквадратичных отклонений наблюдаемых частот от предсказаний уравнения 2 к предсказаниям уравнения 10 составило 4,99 для скворца ( N = 20) и 27,3 для зяблика ( N = 11). . Тест F (Bailey, 1995) показал, что эти отношения дисперсии очень значимы в обоих случаях ( P 0,01), то есть точки на рис.6 значительно ближе к кривым, чем к горизонтальным линиям. Уравнение 10 также предсказывает частоту взмахов крыльев, равную 24.9 Гц по данным Tobalske et al. (Tobalske et al., 1999) для зебрового зяблика ( Taenopygia guttata ), летящего в аэродинамической трубе с долей мощности 0,5, что находится примерно в середине наблюдаемого диапазона частот. Кажется, что частота взмахов крыльев прыгающей птицы действительно реагирует на увеличение силы тяжести, вызванное вариациями доли мощности. Это не результат, который можно было бы предвидеть с помощью эмпирического подхода к влиянию гравитации на частоту взмахов крыльев.
Рис. 6.Наблюдаемые частоты взмахов крыльев у двух ограничивающих видов в зависимости от доли мощности. Кружки и сплошные линии, скворец Sturnus vulgaris ; крестики и пунктир - зяблик Fringilla coelebs . Кривые, прогнозируемая частота взмахов крыльев из уравнения 10; горизонтальные линии, прогнозируемая частота взмахов крыльев для доли мощности 1 (непрерывный взмах).
Массовые работы
Рис.2 и 6 показывают, что частота взмахов крыльев птиц при взмахах и взмахах планерами может быть довольно хорошо предсказана из уравнения 2, в то время как изменение в уравнении 10 расширяет прогноз до ограничения. Уравнение 2, а не уравнение 10, применяется к скольжению с закрылками, потому что сила тяжести не увеличивается в фазе мощности скольжения с закрылками, как при ограничении. Как только частота взмахов крыльев может быть предсказана, то же самое можно сделать и для удельной по массе работы в летных мышцах ( Q m ).Работа с удельной массой - это работа, выполняемая на единицу массы мышцы при каждом сокращении, и она связана с механической мощностью ( P мех ) таким образом:
11
, где f - частота взмахов крыльев, а m musc - оценка массы сократительного компонента летательных мышц. Уравнение 11 применяется к непрерывному взмаху, скольжению закрылка или ограничению. Доля мощности q появляется в знаменателе уравнения 11, потому что P mech - это средняя скорость выполнения работы за весь цикл прыжка или скольжение закрылка, но работа выполняется только во время фазы мощности.
Работа, ориентированная на массу, является хорошей переменной для определения пределов мышечной работоспособности, поскольку она имеет четко определенный верхний предел, который вряд ли будет сильно варьироваться от одного животного к другому. Это не эмпирический результат, а тот, который непосредственно следует из основных свойств механизма скользящей нити, которые были поняты в течение многих лет (Huxley, 1957; McMahon, 1984). Работа с удельной массой пропорциональна произведению напряжения ( s ) и напряжения ( e ) в сократительных белках, в то время как мышца укорачивается (Pennycuick and Rezende, 1984):
12
, где ρ musc - плотность мускулов, обычно принимаемая равной 1060 кг · м –3 .Напряжение имеет верхний предел, который в конечном итоге можно проследить до силы, которая может быть приложена единственной миозиновой нитью (Pennycuick, 1992; Pennycuick, 1998). Штамм также имеет верхний предел, связанный с количеством перекрытий, которое возможно между актиновыми и миозиновыми филаментами (White and Thorson, 1975). Эти верхние пределы могут различаться для разных типов мышц, но вряд ли сильно различаются между мышцами, которые так же похожи, как летательные мышцы разных птиц. Их фактические значения сложно измерить, но это не влияет на связь с удельной массой работы.Например, если мы предположим «наиболее вероятные» значения 240 кН · м –2 для максимального значения напряжения при укорачивании мышцы (не изометрическое напряжение) и 0,20 для максимального напряжения, то уравнение 12 дает максимум значение примерно 45 Дж кг –1 для удельной работы. Хотя это значение, возможно, придется пересмотреть в свете будущих экспериментов, его можно временно использовать для оценки известной производительности некоторых крупных птиц. Например, Flight.bas оценивает, что удельная работа по массе должна составить около 36 Дж кг –1 для очень большого лебедя-кликуна ( Cygnus cygnus ), измерения которого были даны Pennycuick et al.(Pennycuick et al., 1996a), чтобы лететь на V mp . За этим конкретным лебедем наблюдали со спутника, когда он мигрировал из Шотландии в Исландию, и действительно, похоже, он ограничивался скоростью ниже примерно 1,3 V mp , время от времени приземляясь на воду, как и другие лебеди с острова. такое же население.
Максимальная масса тела при работе
Hedenström и Alerstam (Hedenström and Alerstam, 1992) также наблюдали скорости в районе своей оценки V mp для мигрирующих лебедей-шиповников ( Cygnus olor ) в сочетании с минимальной скоростью набора высоты, и приписали этот маргинальный полет производительность ограничена мышечной силой из-за большого размера лебедей в целом.Сочетание нескольких известных тенденций приводит к тому, что удельная работа, необходимая для полета на V mp , у крупных птиц выше, чем у мелких. Механическая сила, необходимая для полета, увеличивается более круто, чем прямо пропорционально массе тела, и можно было бы ожидать, что фракция летных мышц увеличится у более крупных птиц, но, по-видимому, этого не происходит (Greenewalt, 1962). Это означает, что удельная мощность должна увеличиваться с увеличением массы. Удельная работа увеличивается сильнее, чем удельная мощность, потому что частота взмахов крыльев уменьшается у более крупных птиц.Оценивая удельную работу по массе для 150 видов, для которых были доступны надежные (т.е. из первых рук) измерения крыльев, Pennycuick (Pennycuick, 1996b) обнаружил сильную положительную тенденцию для конкретной работы, необходимой для полета на V mp , что варьировались примерно с 0,26-кратной массой тела. Верхний предел для Q м составлял около 48 Дж кг –1 , но эта оценка может быть завышенной, потому что для коэффициента лобового сопротивления использовались чрезмерно высокие значения, которые теперь считаются устаревшими (см. Выше).
Диапазон скоростей
Одна из причин тенденции к более высоким значениям удельной работы у более крупных птиц заключается в том, что V mp у крупной птицы выше, чем у мелкой, при прочих равных условиях. Это означает, что большой птице легче двигаться против встречного ветра, чем маленькой. Лебеди, отслеживаемые Pennycuick et al. (Pennycuick et al., 1996a) наблюдались полеты против ветра до 20 м с –1 , что потребовало бы полета лебедя на скорости 1.1 или 1,2 раза V mp , тогда как зяблик, летящий на 20 м с –1 , будет лететь со скоростью 2,6 V mp (Таблица 2). «Диапазон скоростей», определяемый как отношение максимальной горизонтальной скорости к V mp , может оказаться более 3 у зяблика (см. Выше), но, вероятно, не больше 1,5 у любого мигрирующего лебедя. Обладая примерно такой же долей мускулов, что и у более крупных птиц, у мелких птиц достаточно «запасных» мускулов, чтобы летать с высокими относительными скоростями, когда это необходимо при встречном ветре.
И наоборот, им требуется только часть доступной им мышечной силы, чтобы летать на скоростях около V mp . Когда зяблик не обязан проникать против встречного ветра, удельная работа, требуемая от его мускулов полета, очень мала. По данным Flight.bas, она была бы ниже 10 Дж кг –1 на скорости до 15 мс –1 у зяблика, который непрерывно махал руками (рис. 7). Это приведет к неэффективному преобразованию энергии топлива в работу по причине, показанной на рис.8, которая представляет собой кривую эффективности преобразования энергии АТФ в работу в зависимости от напряжения, рассчитанную на основе теории скользящей нити Хаксли (Huxley, 1957), представленной Мак-Магоном (McMahon, 1984). Левый конец кривой отражает очевидный факт, что если напряжение равно нулю, мышца потребляет энергию топлива, но не работает, и, следовательно, эффективность также равна нулю. Напряжение должно быть достаточно высоким, чтобы не попадать в круто поднимающуюся часть на левом конце кривой, но не настолько высоким, чтобы достигать круто падающей части на правом конце.Согласно рис. 8, удельная работа должна составлять приблизительно от 10 до 48 Дж · кг –1 , чтобы эффективность преобразования была выше 80% от максимального значения. Хотя эти значения подвержены некоторой неопределенности (Pennycuick, 1991), верхний предел, показанный на рисунке, довольно хорошо согласуется с конкретной работой, рассчитанной Flight.bas для крупных птиц, летящих на V mp , и это означает, что маленькие птицы будут необходимо предпринять некоторые действия, чтобы удельная работа оставалась значительно выше 10 Дж кг –1 .
Рис. 7.Расчетные кривые для зяблика Fringilla coelebs массо-удельной работы в сократительной составляющей летных мышц для разных значений доли мощности ( q ). Горизонтальные пунктирные линии, оцененные верхняя и нижняя границы для 80% эффективности преобразования энергии АТФ в работу, предполагая изометрическое напряжение 300 кПа и деформацию 0,2.
Рис. 8.Теоретическая кривая эффективности преобразования энергии АТФ в работу в зависимости от напряжения , нормированного делением на изометрическое напряжение (Pennycuick, 1991).Вертикальные пунктирные линии, верхний и нижний пределы относительного напряжения для коэффициента преобразования 80% или более от максимума. Высокомасштабная работа с удельной массой при изометрическом напряжении 300 кПа и деформации 0,2.
Граничная и рабочая частота
На ум приходят три возможных стратегии для небольшой птицы, летающей рядом с V mp , чтобы увеличить удельную работу ее летающих мускулов без увеличения мощности. Во-первых, птица может задействовать только часть волокон летательных мышц и продолжать непрерывно взмахивать руками.Во-вторых, он может использовать все волокна, кроме скольжения лоскута, поэтому удельная работа увеличивается обратно пропорционально доле мощности. В-третьих, он может использовать все волокна, но прибегать к ограничению, и в этом случае уменьшение доли мощности также увеличивает удельную работу, но более сильно, чем при скольжении закрылками (рис. 7). Однако на рис. 5 показано, что ограничение влечет за собой увеличение мощности и снижение подъемной силы к сопротивлению, что должно быть компенсировано любым увеличением эффективности.
Хотя рис.7 показывает, что ограничение является высокоэффективным методом регулировки конкретной работы в широком диапазоне, рис. 5 показывает, что ограничение влечет за собой штрафы как в мощности, так и в эффективном отношении подъемной силы к сопротивлению, тогда как два других метода этого не делают. Однако ограничение увеличивает частоту взмахов крыльев, и это позволяет мускулам полета адаптироваться к более высокой «рабочей частоте», как это определено Пенникуиком и Резенде (Pennycuick and Rezende, 1984). Это означает, что максимальная скорость деформации миофибрилл может быть установлена на более высокое значение, чем это было бы возможно, если бы мышцы были адаптированы для эффективной работы при непрерывном хлопании.Эффект состоит в том, что для полета на максимальной скорости требуется меньше мышц или, альтернативно, данное количество мышц может справиться с более широким диапазоном скоростей без необходимости работать с чрезмерно низкими значениями удельной работы на низких скоростях. Конечно, остается возможность, что зяблики, летающие в районе V mr , могут задействовать только часть летных мышц, помимо увеличения удельной работы за счет прыжков.
Оптимальная миграция
Любая из альтернативных гипотез, предложенных Hedenström и Alerstam (Hedenström and Alerstam, 1995), предсказывала бы, что птицы «должны» мигрировать с большей скоростью, чем наблюдалось в этом исследовании, за исключением, возможно, случая двух маленьких воробьиных, которые пролетали мимо ограничиваясь.Даже они летели намного быстрее, чем V mp , когда летели против встречного ветра. Прежде чем можно будет проверить какие-либо теории оптимальной миграции, необходимо прежде всего установить, являются ли указанные здесь низкие относительные скорости артефактом из-за ограничения наблюдений только низколетящими птицами. Чтобы проверить это, необходимо измерить скорость птиц на нормальных высотах для миграции на большие расстояния по тому же эталону ( V mp ). Это потребует отслеживания и идентификации высоко летающих птиц, что представляет некоторые технические трудности.Если их удастся преодолеть, и окажется, что крейсерская скорость, наблюдаемая на больших высотах, по-прежнему близка к V mp по расчетам Flight.bas, следующий вопрос, который необходимо рассмотреть, заключается в том, постоянно ли Flight.bas завышает значение V Т.пл. . В настоящее время это кажется маловероятным ввиду экспериментов в аэродинамической трубе Pennycuick et al. (Pennycuick et al., 1996b), в котором было получено хорошее согласие между наблюдаемыми и расчетными значениями V mp после пересмотра в сторону понижения аномально высоких значений, ранее принятых для коэффициента сопротивления тела.Прогнозируемое значение V mp необходимо протестировать на более широком диапазоне видов, учитывая, что такие измерения чувствительны к качеству окружающей среды в аэродинамической трубе (Pennycuick et al., 1997).
Если будет подтверждено, что мигранты, летящие на больших высотах, совершают крейсерские полеты на более высоких скоростях относительно значения V mp , предсказанного Flight.bas, тогда будет мотив для попытки вычислить дополнительные контрольные скорости, соответствующие к различным оптимальным скоростям, предложенным Хеденстремом и Алерстамом (Hedenström and Alerstam, 1995).Конечно, Flight.bas в его нынешнем виде будет генерировать оценки мощности на любой необходимой скорости. Однако лежащая в основе теория включает упрощения (полностью явные в опубликованной версии), которые удовлетворительны при скоростях около V mp , но экспериментально не проверены на более высоких скоростях и, возможно, должны быть изменены в свете будущих экспериментов. В настоящее время неясно, у каких видов птиц (если таковые имеются) есть либо достаточная механическая мощность, доступная от их мускулов, чтобы летать на скоростях, намного превышающих V mp , либо достаточная аэробная способность для длительного плавания на таких высоких скоростях, особенно на больших высотах.Это должно быть установлено путем измерения в высококачественных аэродинамических трубах как выходной механической мощности мускулов полета, так и показателей расхода топлива при горизонтальном полете до самых высоких скоростей, которые могут выдерживать разные птицы.
Самые быстрые птицы в мире
Часто рассматриваемые как величественные существа, парящие над головой, птицы бывают всех форм и размеров и могут похвастаться множеством физических возможностей, таких как способность летать или нырять со скоростью, сравнимой с скоростью конкурирующих транспортных средств.Некоторые, например нелетающий страус, могут бегать с головокружительной скоростью, избегая своих естественных хищников. Эти быстрые животные сочетают тягу, сопротивление, вес и подъемную силу, чтобы достичь полета, но их взмах крыльев и размах помогают определить, насколько быстро они смогут взлететь. Это одни из самых быстрых птиц в мире по определенным навыкам, таким как способность нырять, скорость полета и бега, а также взмах крыльев.
Самая быстрая птица (ныряние): сапсан
Сапсан в пикировании.Изображение предоставлено: Smiler99 / Shutterstock.comСоколы-сапсаны могут развивать скорость до 240 миль в час, ныряя или наклоняясь, используя свои длинные заостренные крылья, которые помогают направлять их путь, преследуя свою добычу, часто более мелкую птицу. В 2005 году шестилетний сокол-сапсан по имени Who Frightful в Соединенных Штатах установил рекорд самого быстрого прыжка среди птиц со скоростью почти 242 миль в час.
На сегодняшний день не замечено ни одного другого вида птиц, который мог бы соперничать с сапсаном по скорости пикирования. Скорость достигается за счет взлета на большую высоту и затем крутого наклона. Чтобы защитить свои легкие, птицы разработали носовой покров, который предотвращает попадание опасного воздуха под высоким давлением в их тела.
Несмотря на впечатляющую скорость пикирования, сокол-сапсан не может сравниться с другими птицами при измерении горизонтального полета - фактически, он не входит в десятку самых быстрых птиц в этой категории.
Самая быстрая птица (в горизонтальном полете): сероголовый альбатрос
Сероголовый альбатрос пролетает над морем на острове Южная Георгия в Антарктиде. Изображение предоставлено: MZPHOTO.CZ/Shutterstock.comСедоголовый альбатрос может развивать скорость более 78 миль в час, рекордсмен - 78 миль.9 миль в час в 2004 году. Альбатрос не только способен развивать огромную скорость, он также может сохранять свою скорость в течение длительного времени, при этом рекордная, находящаяся под наблюдением птица сохраняет свою высокую скорость более восьми часов, путешествуя на юг. Гнездо грузин во время антарктического шторма.
Эти быстрые морские птицы кормятся дальше на юг, чем любые другие в своем роде, от мест гнездования в Южной Атлантике до Антарктики, где они используют сильные штормы в регионе, чтобы летать еще быстрее.Их типичная скорость во время кормодобывания составляет 68 миль в час, а большой размах крыльев помогает им сохранять равновесие и идти в ногу.
Самая быстрая птица (бегает по суше): страус
Страус в пустыне. Изображение предоставлено: COULANGES / Shutterstock.comНесмотря на то, что страус нелетающий из-за своего большого размера, он может развивать впечатляющую скорость по земле и занимает первое место в категории скорости бега среди птиц.Они могут даже обогнать другие виды, достигая максимальной скорости около 45 миль в час пешком, что может выдерживать большие расстояния - страус может преодолеть олимпийский марафон менее чем за 45 минут. Их ноги являются фактором, способствующим их скорости, с шагом от 10 до 15 футов, и достаточно сильны, чтобы убить человека одним быстрым ударом.
Его крылья по-прежнему полезны, хотя страус не отрывается от земли.Имея размах 2,2 метра, крылья помогают страусу уравновесить его и управлять его быстрым бегом с легкими движениями, похожими на крылья самолета. Кроме того, центр тяжести страуса идеально расположен между его широкими крыльями и длинными ногами, что помогает контролировать его баланс независимо от того, с какой скоростью он движется.
Самый быстрый взмах крыльев птицы: колибри с рубиновым горлом
Колибри с рубиновым горлом.Изображение предоставлено: Стив Байлэнд / Shutterstock.comКолибри с рубиновым горлом имеет право хвастаться как птица с самым быстрым взмахом крыльев, особенно во время ухаживания, со скоростью около 200 ударов в секунду. Эта скорость превосходит других колибри, которые обычно совершают около 90 ударов в секунду своими крыльями. Однако во время обычного полета колибри с рубиновым горлом взмахивает крыльями примерно 53 раза в секунду, что помогает ему летать акробатическим способом, с которым немногие другие виды могут соперничать.
Другие Fast Birds
Колибри Анны
Мужчина Колибри Анны.Изображение предоставлено: Keneva Photography / Shutterstock.comКолибри Анны, представитель своего вида среднего размера, обитает на побережье Северной Америки и также может похвастаться средней скоростью полета 61 миль в час. Наиболее впечатляющими являются ритуалы ухаживания самца колибри Анны, который на большой скорости падает на землю, чтобы произвести впечатление на потенциальных партнеров. За каждую секунду своего спуска птица покрывает 385-кратную длину своего тела. Из-за своего крошечного размера (около 2,75 дюйма в длину) ускорение с такой скоростью является самым быстрым воздушным маневром, выполняемым любой птицей - сапсан и его заметное ныряние покрывают всего около 200 длин тела в секунду.
Евразийское хобби
Евразийское хобби принадлежит к семейству соколиных, хотя и относится к меньшей части спектра для этого вида. Он мигрирует на большие расстояния, чтобы провести зиму в Центральной Африке и Южной Азии, достигая максимальной скорости 100 миль в час.Хобби может делать быстрые, подвижные движения в воздухе, используя устойчивый взмах крыльев и долгое скольжение, чтобы поддерживать его скорость. Более быстрые взмахи крыльев помогают увлечению Eurasion быстро ускориться на охоте, перемежаясь с более коротким скольжением. Прыгая за добычей, он расправляет крылья
Кречет
Кречет в полете. Изображение предоставлено Питером Ван Дамом / Shutterstock.comИзвестный как самый большой сокол в мире, кречет известен своими длинными заостренными крыльями, которые помогают при быстром нырянии, что позволяет птице спускаться с большой высоты, чтобы поймать добычу. Во время погружения кречет часто развивает скорость до 130 миль в час, с большим контролем над спуском, используя положение крыльев, что может помочь увеличить или уменьшить скорость и направление при небольшом движении.
Это птица, это самолет
Птицы были источником вдохновения для авиации, поскольку люди пытались имитировать движение крыльев при проектировании первых самолетов.Со временем и развитием технологий эти конструкции изменились, но все еще имели некоторое сходство с движением птиц в полете.
Например, взлет вертолета, который использует свои винты для создания подъемной силы, выталкивая воздух вниз, аналогичен тому, как некоторые птицы полагаются на быстрые взмахи крыльев для достижения того же результата. Обтекаемые формы современных самолетов также похожи на птиц, которые их вдохновляли, с телами, сделанными из легкого материала, схожего с костной структурой низкой плотности у птиц, и заостренным носом, имитирующим птичий клюв.Кроме того, самолеты используют свои крылья и хвост для регулировки наклона, снижения, скорости и направления, как это делают птицы во время длительного полета или ныряния.
10 самых быстрых птиц в мире
Птицы принадлежат к группе эндотермических позвоночных, характеризующихся перьями, беззубыми клювыми челюстями, кладкой яиц с твердой скорлупой, высоким уровнем метаболизма, четырехкамерным сердцем и сильным еще легкий каркас. Страус - это нелетающие птицы девяти футов высотой, в то время как пчелиный колибри - самая маленькая птица длиной два дюйма.
10 самых быстрых птиц в мире
Список 10 самых быстрых птиц в мире с максимальной скоростью в километрах в час и некоторые важные факты приведены ниже:
10. Колибри Анны
Колибри среднего размера, принадлежит к семейству птиц Trochilidae. Встречается на побережье Северной Америки. У него переливающаяся бронзово-зеленая спина, бледно-серая грудь и живот и зеленые бока.
Максимальная скорость: 98.27 км / ч
9. Седоголовый альбатрос
Он также известен как сероголовый моллимавк и принадлежит к семейству альбатросов.
Максимальная скорость: 127 км / ч
8. Голубь (голубь)
Эта птица принадлежит к семейству Columbidae.
Максимальная скорость: 148,9 км / ч
7. Краснозобый крохаль
Эта птица принадлежит к семейству птиц Anatidae.Он встречается в пресноводных озерах и реках северной части Северной Америки, Гренландии, Европы и Азии.
Максимальная скорость: 129 км / ч
6. Шпоновый гусь
Это большая птица из семейства Anatidae, обитающая в водно-болотных угодьях Африки к югу от Сахары.
Максимальная скорость: 142 км / ч
Список основных рыболовных угодий мира
5. Frigatebird
Он принадлежит к семейству птиц Fregatidae и встречается во всех тропических и субтропических океанах.
Максимальная скорость: 153 км / ч
4. Евразийское хобби
Сланцево-серый цвет сверху с темной короной и двумя короткими черными усами, принадлежит к семейству птиц Falconidae. Это дальняя перелетная птица, зимующая в Африке и Азии.
Максимальная скорость: 160 км / ч
3. Игольчатая белогорлая
Он также известен как иглохвостый стриж или иглохвостый стриж.Это перелетная птица, гнездящаяся в Средней Азии и южной Сибири, а также зимующая на юге Индийского субконтинента, Юго-Восточной Азии и Австралии. Он принадлежит к семейству птиц Apodidae.
Максимальная скорость: 169 км / ч
2. Беркут
Это темно-коричневая птица с более светлым золотисто-коричневым затылком оперения. Это одна из самых известных хищных птиц Северного полушария. Он принадлежит к семейству птиц Accipitridae.
Максимальная скорость: 240–320 км / ч
1.Сапсан
Это большой сокол размером с ворону, у него сине-серая спина, белая полость на нижней части тела, а черная голова принадлежит к семейству птиц Falconidae. Он встречается почти везде на Земле, кроме полярного региона.
Максимальная скорость: 389 км / ч
10 самых опасных вод в мире
10 самых быстрых птиц в мире
Название птицы (ранг) | Максимальная скорость |
1.Сапсан | 389 км / ч |
2. Беркут | 240–320 км / ч |
3. Игольчатая белая горловина | 169 км / ч |
4. Евразийское хобби | 160 км / ч |
5. Frigatebird | 153 км / ч |
6.Шпоновый гусь | 142 км / ч |
7. Краснозобый крохаль | 129 км / ч |
8. Голубь (голубь) | 148,9 км / ч |
9. Сероголовый альбатрос | 127 км / ч |
10. Колибри Анны | 98.27 км / ч |
Список вымерших видов морских рептилий
Скорость миграции для выбранной выборки видов птиц.
Голуби были предметом исследований в прошлом, но полученные знания являются неполными и должны быть расширены. Целью исследования было предоставить информацию о различиях в весе и размерах туши, составе туши, приблизительном химическом составе, кислотности, электропроводности, цветовых характеристиках, текстуре, реологических свойствах и микроструктуре мяса, а также некоторых биометрических характеристиках пищеварительной системы. системы у голубей-носителей и королевских голубей, а также для определения того, соответствуют ли две сравниваемые породы ожиданиям потребителей мяса голубей в одинаковой степени.В исследовании участвовали 40 туш почтовых голубей и королевских голубей после трех репродуктивных сезонов. Химический состав был определен с помощью спектроскопии пропускания в ближней инфракрасной области (NIT), цветовых координат в соответствии с CIELab, текстуры в соответствии с анализом профиля текстуры (TPA) и тестов Warner – Bratzler (WB), а реологические свойства мяса в соответствии с релаксацией. тест. Сравниваемые группы голубей значительно различались (p <0,05) по весу и размерам туши, составу тушки (за исключением процента грудных мышц), химическому составу (за исключением содержания коллагена в мышцах ног) и электропроводности, легкости (L *), желтизне (b *). , цветность (C *) и угол оттенка (h *), текстурные характеристики (кроме когезии и силы сдвига Уорнера-Братцлера), реологические свойства, микроструктура большой грудной мышцы, а также общая длина кишечника и его сегментов, двенадцатиперстной кишки диаметр, вес преджелудка, желудка, печени, сердца и селезенки.Пол птиц оказывал значимое (p <0,05) влияние на вес тушки, окружность грудной клетки, процент шеи тушки, содержание коллагена в грудных мышцах и длину слепой кишки. Генотип по половому взаимодействию был значимым (p <0,05) для содержания жира, содержания коллагена, твердости, суммы модулей упругости и суммы модулей вязкости большой грудной мышцы, содержания белка и коллагена в мышцах ног, длины двенадцатиперстной и слепой кишки, тощей кишки. диаметр подвздошной кишки и вес селезенки. Полученные результаты показывают значительное влияние генетического происхождения и пола на пищевую и технологическую ценность мяса, а также на развитие пищеварительной системы голубей.
Как быстро птицы могут мигрировать? на JSTOR
Миграция птиц обычно характеризуется периодами полета, когда топливо израсходовано, и промежуточными периодами остановки, когда топливо складывается. Результирующая общая скорость миграции может быть рассчитана на основе скорости полета, скорости отложения топлива и потребляемой мощности во время полета. Скорость депонирования энергии $ (P _ {{\ rm dep}}) $ может быть оценена как разница между объемом метаболизма и полевой скоростью метаболизма во время остановки.{-0.19}) $, а скорость миграции увеличивается с увеличением массы для парящего полета. Для махового полета скорость и мощность миграции могут быть рассчитаны в соответствии с аэродинамической теорией, а в парящем полете скорость по пересеченной местности может быть оценена по скорости набора высоты (при круговом полете в термиках) и глиссаде. Предполагается, что мощность в парящем / планирующем полете является постоянной кратной базовой скорости метаболизма (BMR). Мы использовали этот подход для расчета ожидаемой скорости миграции для 15 видов. Для небольшой птицы с типичной скоростью выделения энергии $ (P _ {{\ rm dep}} = 1.0 \ cdot {\ rm BMR}) $ и при использовании колебательного полета прогнозируемая скорость миграции составляет около 200 км / день, а максимальная скорость выделения энергии $ (P _ {{\ rm dep}} = 2,5 \ cdot {\ rm BMR} ) $ дает скорость миграции 300-400 км / сутки. У более крупных птиц скорость миграции будет ниже: от 70 до 100 км / день для типичных скоростей депонирования энергии до 150-200 км / день для максимальных скоростей депонирования энергии. У крупных видов парящий полет дает более высокую скорость миграции, чем взмах. Для крупных видов, использующих взмахи крыльев, скорость миграции сама по себе может ограничивать потенциальное расстояние миграции, которое можно учесть в рамках ежегодной программы размножения-миграции.
Journal of Avian Biology публикует эмпирические и теоретические исследования во всех областях орнитологии с акцентом на поведенческую экологию и эволюцию. Желательно, чтобы статьи проверяли гипотезы или исследовали новую почву. Чисто описательные исследования принимаются только тогда, когда они содержат важную информацию, которая вызывает новые вопросы или открывает новые возможности.
Wiley - глобальный поставщик контента и решений для рабочих процессов с поддержкой контента в областях научных, технических, медицинских и научных исследований; профессиональное развитие; и образование.Наши основные предприятия выпускают научные, технические, медицинские и научные журналы, справочники, книги, услуги баз данных и рекламу; профессиональные книги, продукты по подписке, услуги по сертификации и обучению и онлайн-приложения; образовательный контент и услуги, включая интегрированные онлайн-ресурсы для преподавания и обучения для студентов и аспирантов, а также для учащихся на протяжении всей жизни. Основанная в 1807 году компания John Wiley & Sons, Inc. уже более 200 лет является ценным источником информации и понимания, помогая людям во всем мире удовлетворять свои потребности и воплощать в жизнь их чаяния.Wiley опубликовал работы более 450 лауреатов Нобелевской премии во всех категориях: литература, экономика, физиология и медицина, физика, химия и мир. Wiley поддерживает партнерские отношения со многими ведущими мировыми сообществами и ежегодно издает более 1500 рецензируемых журналов и более 1500 новых книг в печатном виде и в Интернете, а также базы данных, основные справочные материалы и лабораторные протоколы по предметам STMS.