
Домовый воробей
Домо́вый воробе́й (лат. Passer domesticus) — наиболее распространённый вид семейства воробьиные. Отряд: Воробьинообразные. Класс: Птицы. Тип: Хордовые. Царство: Животные.
Описание. Длина тела составляет 14–18 см, масса — 21–37 г. Общая окраска оперения — сверху коричневато-бурая, ржавчинного цвета с чёрными пятнами, снизу беловатая или серая. Щёки белые, ушная область бледно-серая. Крылья с желтовато-белой поперечной полосой. Самец отличается от самки наличием большого чёрного пятна («галстука»), охватывающего подбородок, горло, зоб и верхнюю часть груди, а также тёмно-серым (а не тёмно-бурым) верхом головы. У самки голова и горло серые, а над глазом имеется бледная серо-жёлтая полоса
Голос – хорошо известное всем чириканье, которое в разных вариантах используется во многих ситуациях, в т. ч. и в качестве песни. Голос домового воробья пение крики (onbird.
Образ жизни. Почти повсеместно является оседлой птицей. Гнездится воробей отдельными парами, но иногда и колониями, поселяясь непосредственно у жилья человека или вблизи его поселений. На юге ареала нередко строит гнёзда в стороне от них — в древесных и кустарниковых насаждениях, в оврагах, по крутым глинистым обрывам по соседству с полями. Гнёзда устраивает в разнообразных местах — в щелях строений, в норах глинистых и меловых оврагов, в стенках гнёзд крупных птиц (цапель, аистов, орлов), в дуплах деревьев, может занимать скворечники и норы береговых ласточек. Формально относится к категории птиц-дуплогнёздников. На юге нередко сооружает гнёзда открыто, на ветвях различных деревьев.
В средней полосе России предбрачное оживление начинается в марте, иногда в феврале, и сопровождается криками и драками. Во второй половине марта птицы разбиваются на пары и приступают к постройке гнёзд; откладывание яиц происходит в апреле.
Кладка состоит из 4–10, чаще 5–7, белых яиц с буроватыми крапинками и пятнами, насиживание которых занимает 11–13 дней. Птенцов выкармливают самец и самка, преимущественно насекомыми. Через 10 дней после вылупления (в средней полосе в конце мая — начале июня) птенцы вылетают из гнёзда.
Птенцов выкармливают самец и самка, преимущественно насекомыми. Через 10 дней после вылупления (в средней полосе в конце мая — начале июня) птенцы вылетают из гнёзда.
Питается в основном растительной пищей, весной и летом частично насекомыми, которыми также вскармливает птенцов. В рацион воробья входят семена сельскохозяйственных культур, отбросы различных продуктов, которые он подбирает в человеческих поселениях, хлебные злаки в полях, ягоды вишни, смородины, винограда в садах, весной цветочные почки. При отсутствии поблизости полей вылетает кормиться на луга, опушки лесов и в степи, где собирает семена дикорастущих трав и иногда ловит насекомых. На полях часто кормится в смешанных стаях с полевым воробьём, в Средней Азии также с черногрудым.
Места обитания. Будучи синантропным видом — постоянным сожителем человека, домовый воробей хорошо приспособлен к жизни в обстановке, меняющейся под воздействием хозяйственной деятельности человека. Воробья равно можно встретить в сельской и городской местности, в поселениях Крайнего Севера и Средней Азии.
Ареал. Прежде область обитания домового воробья ограничивалась Северной Европой. Впоследствии широко распространился в Европе и Азии (за исключением Арктики, северо-восточных, юго-восточных и центральных районов Азии), а также в Севернойи Восточной Африке, Сенегале, Малой Азии, на Аравийском полуострове и острове Ява. В Италии обитает близкий вид — итальянский воробей (Passer italiae).
Начиная с XX века был завезён в разные страны, широко там расселился и в настоящее время, кроме указанных выше мест, обитает также в Южной Африке, Австралии, Новой Зеландии, Северной и Южной Америке и на многих островах.
Встречается в заповедниках «Хакасский», «Астраханский», «Центральносибирский», «Малая Сосьва», в национальных парках «Шорский», «Нижняя Кама», «Тебердинский», «Марий Чодра».
Охранный статус. В Красном списке Международного союза охраны природы внесен в категорию видов, вызывающих наименьшие опасения.
Лучшее время для наблюдения. Круглогодично, в дневное время.
Использованные источники:
https://ru.wikipedia.org/wiki/%D0%94%D0%BE%D0%BC%D0%BE%D0%B2%D1%8B%D0%B9_%D0%B2%D0%BE%D1%80%D0%BE%D0%B1%D0%B5%D0%B9
https://ptici.info/ptici/vorobey-domoviy.html
http://www.ebirds.ru/vid/387.htm
Полевой воробей - Воробьиные | Некоммерческий учебно-познавательный интернет-портал Зоогалактика
Описание
Полевой воробей (Passer montanus) не имеет такого ярко выраженного полового диморфизма, как его домовый собрат, - самцы и самки этих птиц окрашены практически идентично. Кроме того, он значительно меньше домового воробья: его масса колеблется от 20 до 30 г, тогда как масса домового воробья - от 28 до 38 г. У полевого воробья верх головы, так называемая "шапочка", приятного коричневого цвета, поэтому в западных странах его называют красноголовым воробьем. "Уздечка", полоса под глазом, горло и кроющие уха у него черные, на белых щеках черные точки – по этому признаку полевого воробья легко отличить от домового. Оперение спины, крыльев и хвоста у полевого воробья бурое, часто с темными настволиями и светлыми охристыми окаймлениями перьев; брюшко беловатое, бока шеи также белого цвета клюв зимой буровато-черный с желтым основанием, летом – черный; ноги бледно-бурые. Оперение молодых птиц значительно тусклее, чем у взрослых, верх головы и спина у них серовато-бурые с темными пестринами; брюшко грязно-белое; горло, "уздечка" и кроющие уха серые. Орнитологи различают от 7 до 33 подвидов полевого воробья.
"Уздечка", полоса под глазом, горло и кроющие уха у него черные, на белых щеках черные точки – по этому признаку полевого воробья легко отличить от домового. Оперение спины, крыльев и хвоста у полевого воробья бурое, часто с темными настволиями и светлыми охристыми окаймлениями перьев; брюшко беловатое, бока шеи также белого цвета клюв зимой буровато-черный с желтым основанием, летом – черный; ноги бледно-бурые. Оперение молодых птиц значительно тусклее, чем у взрослых, верх головы и спина у них серовато-бурые с темными пестринами; брюшко грязно-белое; горло, "уздечка" и кроющие уха серые. Орнитологи различают от 7 до 33 подвидов полевого воробья.
Распространение
Как и домовый, полевой воробей распространен исключительно широко: его ареал занимает практически всю Евразию, за исключением районов Крайнего Севера. В большинстве районов нашей страны оба вида воробьев обитают совместно, и лишь на юге Дальнего Востока полевой воробей живет один. Во Владивостоке, Находке и других городах и населенных пунктах Приморья (за исключением Уссурийска, где встречается и домовый воробей) полевой воробей заменяет своего более крупного родственника. Оба вида воробьёв расселились так широко только благодаря большой экологической пластичности. Они сумели прекрасно приспособиться к жизни по соседству с человеком: здесь воробьи имеют разнообразную пищу, места для гнездования и защиту от многих хищников. Интересно, что человек сам способствовал расселению воробьев. Так, полевые воробьи были завезены и выпущены на некоторых островах Индо-Австралийского архипелага, на юге Австралии и в Северной Америке, где единственное пока их местообитание находится в междуречье Миссисипи и Миссури.
Оба вида воробьёв расселились так широко только благодаря большой экологической пластичности. Они сумели прекрасно приспособиться к жизни по соседству с человеком: здесь воробьи имеют разнообразную пищу, места для гнездования и защиту от многих хищников. Интересно, что человек сам способствовал расселению воробьев. Так, полевые воробьи были завезены и выпущены на некоторых островах Индо-Австралийского архипелага, на юге Австралии и в Северной Америке, где единственное пока их местообитание находится в междуречье Миссисипи и Миссури.
Полевой воробей обитает в различных биотопах на севере и в средней полосе, предпочитая редколесья, рощи, кустарники, парки, сады, окраины больших городов. Он избегает селиться лишь в больших сплошных лесах и тайге, куда проникает лишь в поселения человека. В степной полосе полевой воробей живет по обрывам в норах различных птиц, в пойменных лесах, а также в жилье человека. Особенно многочислен он в местах, где имеются поля с зерновыми культурами.
Образ жизни и гнездо
Обычно полевые воробьи гнездятся парами - это типичные моногамы, как и большинство других воробьев. Самец и самка сохраняют верность друг другу в течение всего периода гнездования, а возможно, и всей жизни. Нередко встречаются небольшие колонии полевых воробьев по 5-6 гнезд и более.
Гнезда у воробьев типичные для ткачиковых птиц - шарообразной формы, до 125 мм в диаметре и сделанные очень тщательно. Постройка гнезда занимает много времени, иногда около месяца; оно сплетается из тонких веточек, стеблей различных растений - чаще злаковых трав. Входное отверстие находится сбоку, в верхней части гнезда, его края несколько выдаются вперед, образуя небольшую трубку. Лоток выстилается шерстью, волосами, перьями и пухом; в городе в качестве строительного материала порой используются вата, пакля, нитки и другие материалы. Свои гнезда воробьи умудряются помещать в самых различных местах. По разнообразию мест гнездования они держат первенство среди птиц.
Размножение
Кладка полевого воробья может содержать от 2 до 9 яиц (обычно 5-6). Самка откладывает яйца с начала апреля до первой декады июля. Насиживают кладку на протяжении 11-13 дней и выкармливают птенцов оба родителя. Птенцы оперяются в возрасте 12-17 дней.
Питание
Питаются полевые воробьи семенами, в том числе и культурных злаков, однако, птенцов выкармливает исключительно насекомыми. В негнездовое время они держатся стайками в несколько десятков птиц, часто вместе с домовыми воробьями.
Читайте также:
Что вы знаете про воробья? | Блог пользователя nemo | Живой Ангарск
— Чтож так скучно и не интересно-то тут у вас? — спросил Старый Пират возящихся, сопящих, пищащих чумазых и сопливых ЖАшников. Те навострили ушки, подняли глазки и притихли.
— А ну-ка приготовте каждый по десять пальчиков. Будем вспоминать птичек местных, каких знаем, и эти пальчики загибать!
— Трясогузка... — сказала тихо одна Рыженькая девочка с двумя торчащими косичками, жутко покраснела и спряталась за спины других.
 Опрятный мальчик воспитанно тянул вверх ручку...
Опрятный мальчик воспитанно тянул вверх ручку...— Ну? — строго спрсил Пират.
Мальчик поправил свои штанишки на лямках, встал по стойке "смирно" и пискляво доложил:
— Дятел Желна и Поползень!
Все почтительно замолчали.
— Молодец, М4! — порадовался Пират — Ты всегда был отличником и любителем-краеведом! — А что вы знаете про нашего простого Воробья?
— Он у наших курочек ворует комбикорм!
— И всё? Да-а-а... Садитесь тогда поудобнее, расскажу вам про эту птичку...
Счастливчики забрались Пирату на коленки, остальные сели за стол. Неудачники разместились на лавочке вдоль печки...
Старый Пират NEMO закурил свою трубку и начал:
— Воробей – это одна из самых известных птиц, обитающих по соседству с жилищем человека... — важно сказал он, показывая слайды на своей старой мобилке:
— В России встречаются два вида воробьев: воробей домовой, или городской, и воробей полевой, или деревенский...
Вес воробья всего от 20 до 35 граммов. Между тем, воробей принадлежит к отряду воробьинообразных, в который помимо него входят более 5000 видов птиц. Самым крупным представителем отряда является ворон (его масса около полутора килограмм), самым маленьким – королек (масса до 10 граммов).
Между тем, воробей принадлежит к отряду воробьинообразных, в который помимо него входят более 5000 видов птиц. Самым крупным представителем отряда является ворон (его масса около полутора килограмм), самым маленьким – королек (масса до 10 граммов).
Самец воробья отличается от самки наличием большого чёрного пятна, охватывающего подбородок, горло, зоб и верхнюю часть груди, а также тёмно-серым (а не тёмно-бурым) верхом головы.
Строение глаз воробья таково, что птицы видят мир в розоватом цвете.
Сердце воробья делает до 850 ударов в минуту в состоянии покоя, а во время полета до 1000 ударов в минуту. При этом, сильный испуг чреват для птицы даже смертельным исходом, так как существенно повышает кровяное давление.
Температура тела воробья около 40 градусов.
Воробей за день тратит очень много энергии и поэтому не может голодать более двух суток.
По ряду наблюдений, воробьи образуют пары фактически на всю жизнь.
Жизненный цикл воробья имеет небольшую протяженность, хотя отмечались случаи и 9-ти, и 11-ти летних воробьёв, большинство их не доживает и до 4 лет. Много молодых птиц гибнет ещё в первую зиму, так что средняя продолжительность жизни воробьёв составляет 9 - 21 месяц.
Много молодых птиц гибнет ещё в первую зиму, так что средняя продолжительность жизни воробьёв составляет 9 - 21 месяц.
Кладка воробья состоит из 4—10, чаще 5—7, белых яиц с буроватыми крапинками и пятнами, насиживание которых занимает 11—13 дней.
Воробьи очень плодовиты и в течение лета в северных районах выводят два а в южных три выводка. Второе откладывание яиц происходит во второй половине июня, вылет птенцов — в июле.
Молодых воробьёв с их жёлтым окрасом вокруг клюва в народе называют «желторотиками».
Через 10 дней после вылупления птенцы воробья вылетают из гнёзда.
Питаются воробьи семенами. Обожают коноплю, подсолнечник, пшеничные зерна, но будут клевать и хлебные крошки.
Но малышей воробьи кормят нежным мясом гусениц, бабочек, и польза воробьев, заключающаяся в уничтожении вредителей в огородах, виноградниках и садах, уравновешивает некоторый вред, приносимый едоками зерна. Выкармливая выводок из четырех-шести птенчиков, родители-воробьи суют в оранжевые рты детворы около тысячи разных насекомых.
Воробьи ведут оседлый образ жизни. Только популяции птиц, проживающие в самых северных частях ареала (например, Белоголовый воробей), на зимовку отправляются в более теплые места. Но перелет они совершают, как правило, не очень далекий – до тысячи километров.
Купаясь в песке воробьи чистят перья и избавляются от паразитов. По народной примете подобная песчаная процедура предвещает дождь.
Известны факты установки памятников воробью, сооружённые человеком в знак благодарности за помощь со стороны этой птицы в борьбе против насекомых-вредителей. Первый такой памятник был воздвигнут в середине XIX века в Бостоне.
Но не всегда воробьям ставили памятники. В 1958 году, в Китае открылась компания, по инициативе кормчего Мао Цзэдуна. Компания была начата из-за проблемы с сельскохозяйственными вредителями. Воробьи были объявлены главными врагами, так как по подсчётам кормчего в год они съедали около 20 - 35 тысяч тонн, выращиваемого на полях риса. Китайцам были даны инструкции пугать птиц, так как воробьи больше 15 минут не могут летать без отдыха. В результате воробьи падали на землю мёртвыми.
В результате воробьи падали на землю мёртвыми.
Почти два миллиарда — столько за год, согласно статистике было уничтожено воробьёв. После этих действий урожай был лучше, но его очень сильно поели саранча и гусеницы, численность которых регулировали воробьи. В результате возник голод в период, которого погибло около 20 миллионов человек, а воробьёв пришлось завозить из соседней страны...
Вечерело...
Потрясенные маленькие слушатели молча расходились по домам.
Корабельный кот Васька задумчиво помешивал в закопченом чугунке гречневую кашу...
Пират читал Брема...
...
Воробей на ветке
Это еще одна интересная игрушка с устойчивым равновесием. Тело и голову воробья вылепи из пластилина. Прекрасный клюв получится из семечка подсолнуха. Вдави его тупым концом. Глаза воробья — спичечные головки, хвост — несколько перышек, ноги — из спичек.
На нижнем конце проволоки, воткнутой в тело воробья, укрепи шарик из пластилина. В тело воробья проволока должна входить позади лапок.
В тело воробья проволока должна входить позади лапок.
При достаточно тяжелом грузе воробей будет отлично сидеть на пальце. А если посадить его на ветку в саду, он будет покачиваться, как живой. Можно сажать его и на новогоднюю елку.
Ну а что будет, если груз легче воробья? Усидит наша птичка или свалится? Когда мы ставили карандаш на острие, то выяснили, что равновесие будет устойчиво, если главная тяжесть находится ниже точки опоры. Теперь главная тяжесть — туловище воробья — выше точки опоры. Значит, бедняга упадет?
Не торопись с ответом. Проверим на опыте. Передвигай пластилиновый шарик вверх и вниз по проволоке. Пытаясь уравновесить воробья при разных положениях легкого груза, ты увидишь, что птичка сидит устойчиво, когда груз сдвинут далеко вниз. А если груз высоко, под самым пальцем, то воробей будет опрокидываться.
Выходит, что устойчивость равновесия зависит не только от массы, но и от положения груза.
Для проверки сделай еще один опыт с пластилиновым воробьем. Попробуй уравновесить его, подперев проволоку ребром линейки. Ты увидишь, что чем выше поднят противовес, тем ближе к воробью то место проволоки, на котором она уравновешивается. Это место называют центром массы.
Попробуй уравновесить его, подперев проволоку ребром линейки. Ты увидишь, что чем выше поднят противовес, тем ближе к воробью то место проволоки, на котором она уравновешивается. Это место называют центром массы.
Не жалея времени на пробы, ты увидишь, что воробей сидит на пальце до тех пор, пока центр массы остается ниже точки опоры. А как только он станет выше, воробей упадет.
Значит, не обязательно, чтобы главная масса была внизу. Важно, чтобы ниже точки опоры был центр массы. Тогда при нарушении равновесия он пойдет вверх, и равновесие будет восстанавливаться.
Домовый воробей (Passer domesticus) — Птицы Европейской части России
Облик и поведение. Мелкая птица плотного телосложения. Длина тела 14–18 см, масса 20–39 г. Одна из самых широко распространённых и многочисленных птиц мировой фауны, почти обязательный обитатель большинства населённых пунктов. Вдали от поселений человека встречается крайне редко.
Описание. Самец и самка отличаются друг от друга очень хорошо, окраска самца в брачном оперении заметно отличается от окраски в свежем (осеннем) наряде. У самца в брачном пере верх головы серый, бока головы от глаз до основания шеи и зашейка насыщенно каштанового цвета. Уздечка чёрная, слегка заходит за глаз. Чуть выше и позади глаза маленькое белое пятно. Щёки и кроющие перья уха беловатые или светло-серые, заметно контрастирующие с остальным оперением головы. Чёрное горло сливается с чёрным верхом груди и образует контрастный «галстук». Спина коричневато-рыжая с продольными чёрными полосами. Надхвостье шиферно-серое. Низ грязно-белый, с сероватым или буроватым оттенком. Маховые перья и их кроющие темно-коричневые, с контрастной охристой каймой. Концы средних кроющих второстепенных маховых перьев с широкими белыми кончиками, которые сливаются в заметную белую полоску у сгиба крыла. Перья хвоста коричневато-бурые, с охристыми краями. Глаза тёмно-карие, клюв чёрный, лапы телесного цвета. После осенней линьки, в свежем оперении, самцы выглядят более светлыми и охристыми.
Самец и самка отличаются друг от друга очень хорошо, окраска самца в брачном оперении заметно отличается от окраски в свежем (осеннем) наряде. У самца в брачном пере верх головы серый, бока головы от глаз до основания шеи и зашейка насыщенно каштанового цвета. Уздечка чёрная, слегка заходит за глаз. Чуть выше и позади глаза маленькое белое пятно. Щёки и кроющие перья уха беловатые или светло-серые, заметно контрастирующие с остальным оперением головы. Чёрное горло сливается с чёрным верхом груди и образует контрастный «галстук». Спина коричневато-рыжая с продольными чёрными полосами. Надхвостье шиферно-серое. Низ грязно-белый, с сероватым или буроватым оттенком. Маховые перья и их кроющие темно-коричневые, с контрастной охристой каймой. Концы средних кроющих второстепенных маховых перьев с широкими белыми кончиками, которые сливаются в заметную белую полоску у сгиба крыла. Перья хвоста коричневато-бурые, с охристыми краями. Глаза тёмно-карие, клюв чёрный, лапы телесного цвета. После осенней линьки, в свежем оперении, самцы выглядят более светлыми и охристыми. Серый цвет на голове и надхвостье заменяется серовато-бурым, окраска головы становится менее контрастной. Чёрный цвет на груди исчезает, остаётся только на горле. На перьях крыльев и спины появляются светло-охристые каёмки, низ тела также приобретает светло-охристый оттенок. Клюв меняет окраску с чёрного на бурый, с жёлтым основанием.
Серый цвет на голове и надхвостье заменяется серовато-бурым, окраска головы становится менее контрастной. Чёрный цвет на груди исчезает, остаётся только на горле. На перьях крыльев и спины появляются светло-охристые каёмки, низ тела также приобретает светло-охристый оттенок. Клюв меняет окраску с чёрного на бурый, с жёлтым основанием.
Окраска самки значительно более тусклая и не подвержена столь заметной сезонной изменчивости. Верх головы охристо-бурый, от глаза к зашейку проходит широкая светло-охристая бровь, щёки серовато-охристые, горло беловатое. Окраска верха тела, крыльев и хвоста, как у самца, но более тусклая, без шиферно-серых тонов на голове и надхвостье, светлая полоса на крыле не белая, а рыжевато-охристая. Низ тела беловатый с охристым оттенком. Клюв бурый с жёлтым основанием.
Молодые птицы по окраске очень похожи на самку, но ещё более тусклые, с полным отсутствием в оперении чёрного цвета. Клюв светлее, чем у самки, с жёлтыми краями и жёлтыми углами рта. От полевого самец домового воробья отличается серым верхом головы, отсутствием чёрного пятна на щеке, наличием чёрного цвета не только на горле, но и на верхней части груди. От черногрудого самец домового воробья отличается серым верхом головы, значительно более светлой спиной, отсутствием чёрных пестрин на боках тела и на надхвостье. Самка домового воробья отличается от самки черногрудого воробья отсутствием даже слабых пестрин на боках.
От черногрудого самец домового воробья отличается серым верхом головы, значительно более светлой спиной, отсутствием чёрных пестрин на боках тела и на надхвостье. Самка домового воробья отличается от самки черногрудого воробья отсутствием даже слабых пестрин на боках.
Голос. Звуковой репертуар небогат, состоит из различных по длительности и тембру криков «чиррп», «чррии» или «чиип», употребляемых с разной громкостью и в разных сочетаниях. Оформленной песни нет, вместо неё используются повторяющиеся серии тех же самых криков. При исполнении «песни» самец принимает характерную позу, с опущенными до земли крыльями и задранными вверх головой и хвостом.
Распространение, статус. Интродуцирован на всех материках (за исключением Антарктиды) и большинстве островов. Северные пределы распространения ограничены зоной субарктики, в арктических посёлках гнездится спорадически. На большей части территории России распространён повсеместно, будучи тесно связанным с антропогенными ландшафтами. Является одной из самых многочисленных птиц населённых пунктов и разнообразных антропогенных ландшафтов.
Является одной из самых многочисленных птиц населённых пунктов и разнообразных антропогенных ландшафтов.
Образ жизни. В силу очень тесной привязанности к человеку ведёт почти исключительно оседлый образ жизни. Часто поселяется разрозненными колониями или небольшими поселениями. Предпочитает гнездиться в закрытых полостях и нишах различных построек и сооружений человека. Возможно гнездование в естественных дуплах, дуплянках и норах. В степной зоне на юге региона может гнездиться открыто — в кронах деревьев и кустарников. Строят гнездо оба члена пары. Гнездовая постройка представляет собой рыхлую чашу из сухих стеблей травы, растительных волокон, пуха и шерсти. В случае открытого гнездования гнездо имеет форму большого неаккуратного шара из травы с боковым входом. В кладке 4–6 беловатых с зеленоватым или голубоватым оттенком яиц с мелкими серыми или бурыми крапинками; окраска яиц очень изменчива. Насиживает в основном самка, но иногда её может сменять самец. Насиживание длится 12–14 дней, выкармливают птенцов оба родителя в течение 14–17 дней. В течение лета пара может вырастить два и даже три выводка.
В течение лета пара может вырастить два и даже три выводка.
Основу питания составляют зерна разнообразных злаков, её дополняют различные семена, ягоды и плоды, молодые листья и побеги, насекомые, а также любые съедобные объекты, которые птица может обнаружить в городской среде.
Домовый воробей (Passer domesticus)
Другие виды этого семейства:
Воробей- зимующая птица Якутии | Зоомир
Якутия – самая большая по территории республика в России и самая мощная и вечная мерзлота в мире – от 200 и более тысячи метров, она занимает около половины площади страны и почти всю территорию Якутии.
Зима долгая и лютая, весна многоцветная и бурная, а лето знойное и короткое. Нет другого места на планете, где амплитуда колебаний температуры достигла бы ста градусов: морозы за шестьдесят, зной под сорок. Летние ночи белые и светлее, и долгие петербургских.
Такой суровый климат не пугает птиц и зверей. У нас много разных видов птиц, и зимующих, и перелетных.
У нас много разных видов птиц, и зимующих, и перелетных.
Хочу рассказать о маленьких воробьях, за которыми я наблюдаю и летом, и зимой.
Маленький воробей – гнездящаяся оседлая птица. У самца верх головы серый, горло и верхняя часть груди черные, от клюва через глаз к уху идет коричневая полоса; брюшко и полоски на крыльях беловатые.
У самки общий тон окраски буровато серые; на спине светлые и темные пестрины; нижнее сторона от кончика носа до кончика хвоста 14 см. длина крыла 73-75 мм. Масса воробья около 29-41 гр.
В период гнездования у воробьев много хлопот. Сначала они находят себе пару. В брачный период самцы становятся еще более шумливыми и задиристыми. Они постоянно устраивают драки. В которых каждый соперник старается победить, чтобы отвоевать себе хорошее место для постройки гнезда. То место, которое они выбрали становится их территорией. Например, они строили гнездо под крышей нашего дома. Гнездо строят обе птиц. Строят из стеблей, трав, пакли, шерсти, перьев. В это время не трудно заметить где птицы строят гнездо. По всюду можно увидеть птиц несущих что-нибудь в клюве. У птиц нет зубов.
В это время не трудно заметить где птицы строят гнездо. По всюду можно увидеть птиц несущих что-нибудь в клюве. У птиц нет зубов.
Воробьи имеют две кладки. На конец мая и на конец июня. Кладка состоит из 4-8 сероватых с буроватыми пятнами яиц. Размеры яиц 23х16 мм, масса 2,8-3,4 г. Насиживают обе птиц в течение 11-13 суток. После окончания насиживания из яиц вылупляются птенцы. Птенцы появляются на свет слепыми и голыми. Птенцы громко пищать. Они всегда хотят есть. Широко раскрывают клювы. Родители кладут в них принесенную пищу. И вновь отправляются на поиски. Не так то просто прокормить прожорливых птенцов. В это время я ложу на кормушку черви, маленьких насекомых. Через несколько дней птенцы могут есть и жуков, и комаров. В гнезде они находятся 10-12 суток. После вылета птенцы в течение 5-6 дней докармливаются. Родители учат их летать, искать пищу, спрятаться от врагов.
Воробьи ухаживают за своими перышками, принимают «пылевую ванну», и прихорашиваясь чистят их клювом.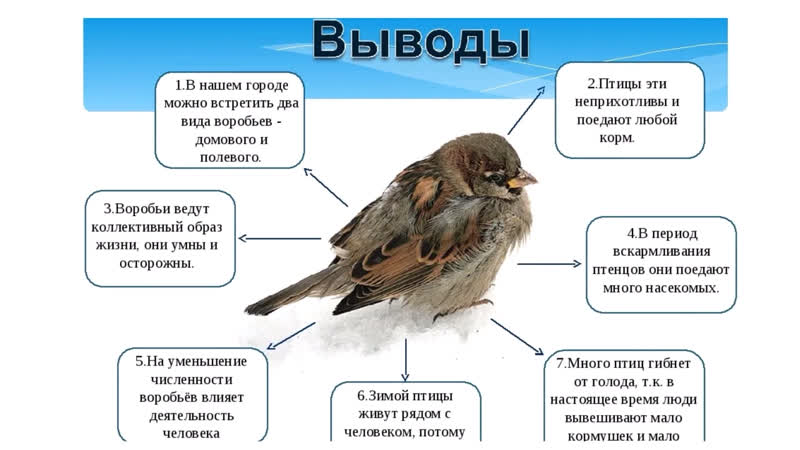
Для маленьких воробьев проживать в наших холодных северных условиях очень трудно. Зимой трудно добывать что-нибудь съестное. Я знаю, что птицы погибают не от холода, а от голода, поэтому я регулярно подкармливаю птиц. Воробьи практически всеядны: они питаются пищевыми отходами, крошками хлеба, семенами, насекомыми. Наша кормушка висит в саду. Там у нас растут бузина, рябина, береза. Воробьи постоянно залетают в сад. Кормлю я их днем. Иногда бросаю семена на землю. Они не боятся меня, но близко не подходят, сидят и наблюдают за мной. Я тихонько отойду и тут они бросаются на еду, их бывает до 50, ходят стаями.
Воробьи вредят посадкам, поедая большое количество семян, но одновременно приносят и пользу, помогая в борьбе с насекомыми – вредителями. Многие виды птиц имеют большое значение в сокращении численности насекомых – вредителей сельского и лесного хозяйства, переносчиков возбудителей опасных заболеваний человека и животных. Особенно много уничтожают насекомых птицы в период выкармливания птенцов. Они выполняют роль санитаров в природе.
Они выполняют роль санитаров в природе.
Все мы должны проявлять заботу о птицах. Птицы – наши друзья.
Похожие статьи
Виды воробьев место обитания внешний вид. Домовой воробей описание питание отличие от полевого воробья
Ещё со школы помню, что у нас водится минимум два вида воробьёв: воробей домовый и воробей полевой. Но в чём их отличие — забыл совершенно. А тут как-то шёл с фотоаппаратом, а у кормушки на кустах копошилась стайка воробьёв. Поснимав их портреты, решил более подробно разобраться с вопросом воробьиной систематики.
Воробей полевой (Passer montanus) отличается немного меньшими размерами и некоторой стройностью по сравнению с домовым, имеет хорошо заметные чёрные «серёжки» на белых щеках, и коричневую «шапочку» на голове.
Воробей домовый (Passer domesticus) чуть крупнее, более драчливый, потому полевой воробей предпочитает с ним не связываться. У домового воробья выраженный половой диморфизм — самцы и самки имеют сильно различающуюся окраску (у полевого — одинаковую). Самцы имеют больше коричневых вкраплений, более яркие, а самочки — серые.
Черный «галстук» у полевого воробья выражен слабо, небольшим черным пятном ниже клюва.
У самца домового воробья большое чёрное пятно, охватывающее подбородок, горло, зоб и верхнюю часть груди.
Считается, что домовый воробей к нам пришел из Средиземноморья и Ближнего Востока, в то время как полевой — из Ближней Азии. Домовый, оправдывая свое название постоянно живёт рядом с человеком, и уже успел освоить все широты, а полевой предпочитает в сытное летнее время жить на природе, а в городе перезимовать неблагоприятные условия.
В этот же день сфотографировал на дереве пару белых трясогузок (Motacilla alba), тоже весьма обычных для города птиц. Длинный качающийся хвост (за что она и получила свое название), серенький верх, белый низ, белая голова с чёрным горлом и шапочкой.
Длинный качающийся хвост (за что она и получила свое название), серенький верх, белый низ, белая голова с чёрным горлом и шапочкой.
При том, что охотно живёт рядом с человеком, трясогузка — всё-таки перелетная птица, но прилетающая в наши края очень рано, в самом начале весны.
Подобно своему родственнику домовому, полевой воробей (Passer montanus) - одна из самых распространенных и обычных наших птиц. Далеко не все различают домовых и полевых воробьев по внешнему виду, тем более что нередко они держатся вместе в общих стаях.
Между тем различия у этих видов довольно существенны. Во-первых, полевой воробей не имеет такого ярко выраженного полового диморфизма, как его домовый собрат. Самцы и самочки окрашены совершенно одинаково. Во-вторых, он значительно меньше домового воробья: его масса колеблется от 20 до 30 г, тогда как масса домового воробья - от 28 до 38 г.
Окраска взрослых полевых воробьев нарядна. Верх головы - "шапочка", коричневая. Ввиду этого в западных странах его называют красноголовым воробьем. "Уздечка", полоса под глазом, горло и кроющие уха черные, на белых щеках черные точки. (Это хороший отличительный признак.) Бока шеи также белого цвета. Оперение спины, крыльев и хвоста бурое, часто с темными настволиями и светлыми охристыми окаймлениями перьев. Брюшко беловатое, темнеющее к бокам. Клюв зимой буровато-черный с желтым основанием, летом - черный. Ноги бледно-бурые. Оперение молодых птиц значительно тусклее, чем у взрослых. Верх головы и спина у них серовато-бурые с темными пестринами. Брюшко грязно-белое, горло, "уздечка" и кроющие уха серые. Орнитологи различают от 7 до 33 подвидов полевого воробья.
Ввиду этого в западных странах его называют красноголовым воробьем. "Уздечка", полоса под глазом, горло и кроющие уха черные, на белых щеках черные точки. (Это хороший отличительный признак.) Бока шеи также белого цвета. Оперение спины, крыльев и хвоста бурое, часто с темными настволиями и светлыми охристыми окаймлениями перьев. Брюшко беловатое, темнеющее к бокам. Клюв зимой буровато-черный с желтым основанием, летом - черный. Ноги бледно-бурые. Оперение молодых птиц значительно тусклее, чем у взрослых. Верх головы и спина у них серовато-бурые с темными пестринами. Брюшко грязно-белое, горло, "уздечка" и кроющие уха серые. Орнитологи различают от 7 до 33 подвидов полевого воробья.
Как и домовый, полевой воробей распространен исключительно широко : его ареал занимает практически всю Евразию, за исключением районов Крайнего Севера. В большинстве районов нашей страны оба вида воробьев обитают совместно, и лишь на юге Дальнего Востока полевой воробей живет один. Во Владивостоке, Находке и других городах и населенных пунктах Приморья (за исключением Уссурийска, где встречается и домовый воробей) полевой воробей заменяет своего более крупного родственника.
Как и домовый, полевой воробей получил такое широкое распространение только благодаря большой экологической пластичности. Он сумел хорошо приспособиться к жизни по соседству с человеком. Здесь птицы имеют разнообразную пищу, места для гнездования и наконец надежную защиту от многих хищников. Человек сам способствовал расселению воробьев. Так, полевые воробьи были завезены и выпущены на некоторых островах Индо-Австралийского архипелага, на юге Австралии и в Северной Америке, где единственное пока их местообитание находится в междуречье Миссисипи и Миссури и занимает всего около 22 000 квадратных км.
Полевой воробей обитает в различных биотопах на севере и в средней полосе, предпочитая редколесья, рощи, кустарники, парки, сады, окраины больших городов. Избегает селиться в больших сплошных лесах и тайге, куда проникает лишь в поселения человека. В степной полосе живет по обрывам в норах различных птиц, в пойменных лесах, а также в жилье человека. Многочислен в местах, где имеются поля с зерновыми культурами.
Обычно полевые воробьи гнездятся парами - это типичные моногамы, как и большинство других воробьев. Самец и самочка сохраняют верность друг другу в течение всего периода гнездования, а возможно, и всей жизни. Нередко можно найти и небольшие колонии по 5-6 гнезд и более. Гнезда типично ткачиковые - шарообразной формы, в диаметре до 125 мм, сделаны тщательно, так что постройка гнезда занимает много времени, иногда около месяца. Сплетается оно из тонких веточек, стеблей различных растений - чаще злаковых трав. Входное отверстие находится сбоку, в верхней части гнезда, его края несколько выдаются вперед, образуя небольшую трубку. Лоток выстилается шерстью, перьями и пухом; в городе в качестве строительного материала порой используются вата, пакля, нитки и другие материалы. Свои гнезда воробьи умудряются помещать в самых различных местах. По разнообразию мест гнездования они держат первенство среди птиц. В норах, сделанных птицами (ласточками-береговушками, щурками, каменками) и зверьками (сусликами, хомяками, песчанками), и под крышами строений, в трещинах глинобитных построек, обрывов, скал и в колодцах, в дуплах деревьев и пустотах пней, в старых гнездах мелких птиц и скворечниках, синичниках и других искусственных дуплянках, в основании гнезд некоторых крупных птиц и, наконец, просто на ветвях деревьев.
В настоящее время полевой и домовый воробьи отнесены к категории дуплогнездников. У типичных дуплогнездников яйца не имеют защитной окраски (белые), а у воробьев, напротив, отличаются хорошо заметной пигментацией в виде многочисленных буроватых пятен на светло-оливковом или кремовом фоне.
Воробьи, самые распространенные птицы больших и маленьких городов, деревень и поселков. Люди настолько к ним привыкли, что даже не догадываются о местах возникновения этих крох, которые, между прочим, находятся на другом конце планеты. Родиной домовых воробьев считается Азия, а также Средиземноморье и Ближний Восток. Чтобы узнать об этих птицах другие интересные факты, необходимо выяснить все об их внешнем виде, поведении, питании и привычках.
Описание воробьев
В населенных пунктах можно встретить два вида этих крошечных птичек - домового и полевого. Домовой воробей всегда обитает рядом с человеком. Представители этого вида птиц замечательно адаптировались к условиям проживания вблизи людей. Их не пугает даже жесткий климат, который преобладает в большей части городов России. Воробьи - это оседлые птицы. Они могут откочевывать ближе к югу только из самых холодных городов с лютыми зимами.
Их не пугает даже жесткий климат, который преобладает в большей части городов России. Воробьи - это оседлые птицы. Они могут откочевывать ближе к югу только из самых холодных городов с лютыми зимами.
Внешний вид птиц
Домовой воробей обладает очень небольшими размерами. Длина его тела колеблется от 14 до 18 см. Вес этой маленькой птицы может быть от 25 до 39 г. Голова довольно крупная по отношению к телу, имеет круглую форму. Клюв у воробья широкий, конический. Его длина может достигать 1,5 см. Птица выглядит крепкой и довольно крупной для своего веса. Хвост обычно имеет размер в 5-6 см. Длина конечностей около 2-2,5 см.
Самки воробьев значительно меньше самцов. По половому признаку отличается и цвет их оперения. Верхняя часть тела птицы всегда коричневая. При этом нижняя имеет светло-серую окраску. Крылья воробьев обрамлены бело-желтыми поперечными полосами. Отличия между самкой и самцом заключаются в цвете головы и шеи. У самцов макушка всегда темно-серого оттенка, а область под глазами светло-серая. Горло и грудь обрамляет черное пятно. У самок макушка и шея всегда светло-коричневая. Во время брачного периода цвет перьев у птиц значительно темнеет. Описание домовых воробьев в это время может слегка не совпадать с вышеперечисленными характеристиками.
Горло и грудь обрамляет черное пятно. У самок макушка и шея всегда светло-коричневая. Во время брачного периода цвет перьев у птиц значительно темнеет. Описание домовых воробьев в это время может слегка не совпадать с вышеперечисленными характеристиками.
Размножение воробьев
Домовой вид этих птиц всегда селится около человеческого жилья. Они могут гнездиться отдельными парами, а иногда соединяются в группы. Гнезда воробьи вьют на деревьях, в разломах и щелях старых зданий, под крышами домов, а также в зарослях кустарников. Постройкой жилья всегда занимаются оба представителя пары. Для этого они используют сухую траву, солому, мелкие ветки. По центру гнезда всегда сооружается углубление, чтобы оттуда не выпали яйца или птенцы.
Чаще всего самка начинает делать кладку в апреле. В гнезде после этого может насчитываться до 10 яиц. Они имеют белый цвет и бурые пятнышки. Период инкубации длится примерно 2 недели. Когда птенцы вылупляются, самка и самец начинают вместе кормить их насекомыми. Способность летать малыши обретают уже через две недели после рождения. Продолжительность их жизни может достигать 10 лет. Однако даже до 4 лет доживает лишь малая часть птиц.
Способность летать малыши обретают уже через две недели после рождения. Продолжительность их жизни может достигать 10 лет. Однако даже до 4 лет доживает лишь малая часть птиц.
Чем питаются воробьи
Их рацион в большей части состоит из продуктов растительного происхождения. Домовые воробьи обожают различные семена, зерна злаковых растений и трав. Однако чаще всего им приходится довольствоваться тем, что доступно в населенных пунктах. Потому воробьи поедают отходы продуктов питания человека, насекомых, ягоды и даже почки на деревьях.
Не всем известно, чем питается домовой воробей в зимний период. Ведь в это время из доступа исчезает основная часть рациона этих пернатых, которая состоит из растительности. К счастью, птиц часто подкармливают люди. Всевозможные кормушки с семечками и сухарями могут сохранить жизни сотням птиц, которые не выдерживают голодную зиму и погибают от холода. Как и многим другим пернатым, воробьям необходим песок для хорошего переваривания пищи. Иногда вместо него птицы едят мелкие камешки и засохшие твердые зерна.
Иногда вместо него птицы едят мелкие камешки и засохшие твердые зерна.
Полевые воробьи
Они не настолько привязаны к местам обитания людей. Потому часто оседают в степной и полевой местности. От этой особенности произошло и название вида птиц. Домовой воробей обитает в городах и жилых районах. А вот полевых можно встретить на окраинах поселков, поэтому их часто называют деревенскими. Эти птицы не имеют внешних отличий по половому признаку. Самки и самцы обладают одинаковым цветом перьев и размерами.
Полевые воробьи предпочитают теплый и сухой климат. Они никогда не живут вместе с домовым видом пернатых. Если им приходится пересечься, это всегда сопровождается драками и конкуренцией за территорию. Каждый из видов обладает высокой социальной активностью. Эти пичуги не боятся ни людей, ни домашних животных. Поэтому очень часто можно увидеть, как воробей нагло поедает из миски корм уличной собаки, которая мирно спит, не замечая, что ее объедает маленькая птичка.
Во время весеннего и летнего периода полевые воробьи едят насекомых, а после созревания урожая переходят на пищу с полей, садов и виноградников. Растительного корма в этот период им бывает вполне достаточно. При наступлении холодов птицам приходится довольствоваться зернами и семенами от сорняков. Иногда они залетают во дворы жилых домов, чтобы отыскать там для себя немного пищи.
Отличия полевых и домовых видов воробьев
Далеко не все знают, как отличить полевого воробья от домового. Полевая разновидность птиц немного похожа на самцов домовых пернатых. Но при этом они имеют более изящную форму и меньший вес. Взрослая особь обладает длиной тела от 12 до 14 см. Главное отличие этих видов состоит в цвете макушки и затылка. Эти части тела у них имеют ярко-каштановый окрас. Также в области ушей и под клювом у полевых воробьев находятся небольшие черные пятна. Шею птиц обрамляет воротник из белоснежных перьев, а их крылья имеют не одну, а две светлых полосы.
Оба вида воробьев имеют высокий уровень смертности. Несмотря на то что эти пернатые способны прожить до 10 лет, мало кто из них переживает даже свою первую зиму. Как и все птицы, проживающие в тяжелых климатических условиях и дикой природе, они ежедневно подвергаются различным опасностям. Нехватка пищи в зимний период является главной из них. Именно поэтому люди с наступлением холодов мастерят кормушки из подручных материалов, а затем засыпают их семенами подсолнечника или других растений. Такая доброта и забота со стороны человека ежегодно спасают сотни воробьев от голодной смерти.
Несмотря на то что эти пернатые способны прожить до 10 лет, мало кто из них переживает даже свою первую зиму. Как и все птицы, проживающие в тяжелых климатических условиях и дикой природе, они ежедневно подвергаются различным опасностям. Нехватка пищи в зимний период является главной из них. Именно поэтому люди с наступлением холодов мастерят кормушки из подручных материалов, а затем засыпают их семенами подсолнечника или других растений. Такая доброта и забота со стороны человека ежегодно спасают сотни воробьев от голодной смерти.
Воробей домовой - это самая известная во всем мире птица. Воробей принадлежит к тем немногим видам пернатых, которые стали незаменимыми жителями сельских и городских улиц. Кажется, что без этих шустрых соседей нам было бы уже скучно жить.
Воробей домовой: описание
Воробушек - птичка маленькая, длина его тела - около 15-17 см, вес - 24-35 г, но при этом он имеет крепкое телосложение. Голова округлой формы и довольно большая. Клювик длиной около полтора сантиметра, толстенький, конической формы. Хвостик приблизительно 5-6 см, лапки - 1.5-2.5 см. Самцы по размерам и весу больше самок.
Хвостик приблизительно 5-6 см, лапки - 1.5-2.5 см. Самцы по размерам и весу больше самок.
Окрасом перьев воробьи-девочки и воробьи-мальчики тоже отличаются. Одинаковая у них верхняя часть тела - коричневая, нижняя часть - светло-серая и крылья с бело-желтой полосой, расположенной поперек. Заметное отличие между самками и самцами в окрасе головы и грудки. У мальчиков верх головы темно-серый, ниже глаз - светло-серое оперение, на горлышке и груди хорошо различимое черное пятно. У девочек голова и шея светло-коричневые.
Экология домового воробья
Воробьи обитают рядом с человеческим жильем, они разлетелись на данное время почти по всему миру, но изначально родиной этих пернатых считается большая часть Европы и
Воробей домовой встречается в населенных пунктах, начиная с запада Европы и до берегов на севере Европы достигает арктического побережья, Сибирь тоже населена этими проворными маленькими птичками. На большей части Восточной и Центральной Азии воробей не обитает.
Птицы способны прекрасно адаптироваться в тех условиях, в которых оказались. Это оседлые пернатые, только из северных холодных мест на время морозной зимы перекочевывают туда, где теплее, в южном направлении.
Образ жизни
Как уже говорилось ранее, воробей домовой любит селиться рядом с людьми, может, из-за этого получил название "домовой". Серые пернатые могут жить парами, но бывает, что создают целые колонии. Например, при кормлении всегда собираются в большие стаи. Когда не нужно сидеть в гнездах на яйцах или с на ночь устраиваются в кустарниках или на ветках деревьев.
В воздухе птица развивает скорость полета до 45 км/час, ходить по земле, как большинство других пернатых, воробей не может, передвигается прыжками. В водоеме не утонет, так как умеет плавать, к тому же еще и ныряльщик неплохой.
Размножение
В брачный период домовые воробьи делятся на пары, затем самец и самка вместе приступают к сооружению жилища. Гнезда строят в щелях сооружений и зданий, в дуплах, в норах, на склонах оврагов, в кустарниках и на ветках деревьев. Воробьиный домик делается из маленьких веточек, сухой травы и соломы.
Воробьиный домик делается из маленьких веточек, сухой травы и соломы.
На протяжении апреля будущая мама-воробей откладывает яйца, в гнезде бывает от 4 до 10 яиц, беленьких с бурыми пятнами. Через 14 дней после того, как самка села на яйца, на свет появляются беспомощные птенцы. За вылупившимся потомством папа и мама ухаживают вместе, кормят малышей насекомыми. Уже через две недели птенцы вылетают из гнезда.
Продолжительность жизни
Воробьи в природе живут достаточно долго, продолжительность их жизни - около 10-12 лет. Зафиксирован случай долгожительства - воробей родом из Дании прожил целых 23 года, другой его сородич немножко не дотянул до своего двадцатого дня рождения.
Проблема этих пернатых в том, что очень много гибнет молодых птичек, не доживших до одного года. Самое тяжелое для молодняка время - зима. Если им удастся дожить до своей первой весны, то у них есть шанс встретить старость. На данное время около 70% воробьиной молодежи не доживает до года.
Питание
Воробей домовой вполне может обходиться без воды, необходимое для существования количество влаги он получает из сочных ягодок. Питаются птицы в основном растительной пищей. Любимое лакомство - семена Воробей не переборчивый, кушает то, что попадется, в его рацион входят семена трав, почки деревьев, различные ягоды. Отходами продуктов из мусорных баков эти пернатые тоже не брезгуют, опыт им подсказывает, что в этих железных коробках можно найти много чего вкусненького. Насекомые в воробьиное меню попадают редко, только в период выкармливания птенцов жучки и червячки становятся ежедневной пищей, так как именно ими птицы-родители кормят своих детенышей. Не забывают воробушки также и о песке, он необходим для птичьего желудочка, чтобы переваривать пищу. Если песком разжиться не получается, то используются маленькие камушки.
Подсемейство Воробьи настоящие
К подсемейству Воробьи настоящие относятся домовой воробей, снежный вьюрок, полевой воробей. Хочется обратить внимание на снежного вьюрка, в народе называемого снежный воробей.:max_bytes(150000):strip_icc()/house-sparrow-female-5a11f1dc4e4f7d00366e6627.jpg) Эти птицы довольно красивые, они светлее и крупнее домового. Сверху снежный вьюрок серовато-бурый, а снизу белый, крылья черно-белые. Если наблюдать за птицей в полете, то создается видимость белой птицы с черными пятнышками. Горло вьюрка-самца черное, голова серая, хвост длинный белый с расположенной вдоль. Этот вид воробьев назвали «снежный» из-за их почти белого оперения.
Эти птицы довольно красивые, они светлее и крупнее домового. Сверху снежный вьюрок серовато-бурый, а снизу белый, крылья черно-белые. Если наблюдать за птицей в полете, то создается видимость белой птицы с черными пятнышками. Горло вьюрка-самца черное, голова серая, хвост длинный белый с расположенной вдоль. Этот вид воробьев назвали «снежный» из-за их почти белого оперения.
Полевой, в отличие от снежного, намного меньше домового. Воробей полевой и домовой (самцы) похожи между собой окрасом корпуса и крыльев, отличить их можно легко по окрасу головы. Полевой сородич домового «одет» в каштановую шапочку, которая отделяется от коричневатой спинки узеньким беленьким ошейником. На белых щечках полевого воробья посажено по черному пятну, на горлышке пятно совсем маленькое. Самцы и самки этого вида птиц «одеты» в одинаковую одежку, их окрас не отличается ничем.
Как домовой, так и полевой воробьи селятся рядом с людьми. Полевые, это заметно по названию, в большинстве обитают в сельских населенных пунктах, а домовые, соответственно, в большей мере - городские жители. Птицы стараются держаться подальше стая от стаи, смешанные колонии обоих видов встречаются очень редко. Белые, черные, серые - разница между воробьями не слишком большая, накрепко объединяет их одно - соседство с человеком. Жизнь без этих беспокойных птичек уже даже и не представляется, покидать они нас не собираются, так что пернатое соседство нам обеспечено очень надолго.
Птицы стараются держаться подальше стая от стаи, смешанные колонии обоих видов встречаются очень редко. Белые, черные, серые - разница между воробьями не слишком большая, накрепко объединяет их одно - соседство с человеком. Жизнь без этих беспокойных птичек уже даже и не представляется, покидать они нас не собираются, так что пернатое соседство нам обеспечено очень надолго.
Верабей палявы
Вся территория Беларуси
Семейство Воробьиные - Passeridae.
В Беларуси - P. m. montanus.
Обычный гнездящийся, оседлый вид.
Несколько мельче домового воробья, половой диморфизм в окраске не выражен. Верх головы и затылок каштановые, щеки светло-серые с черным пятном посередине, на горле также небольшое черное пятно.
Грудь, брюшко, надхвостье, маховые и рулевые перья буровато-серые, спина бурая с темными продольными пестринами. Клюв темно-серый, ноги бурые. Оперение молодых птиц лишь несколько более тусклое,
чем взрослых. Вес самца 19,5-27,5 г, самки 19-26 г. Длина тела (оба пола) 14-15 см, размах крыльев 20-22 см. Длина крыла самцов 7-7,5 см, хвоста 5,5-6 см, цевки 1,5-2 см, клюва 0,9-1,2 см. Длина
крыла самок 6-7,5 см, хвоста 5-6 см, цевки 1,5-2 см, клюва 0,9-1,2 см.
Длина тела (оба пола) 14-15 см, размах крыльев 20-22 см. Длина крыла самцов 7-7,5 см, хвоста 5,5-6 см, цевки 1,5-2 см, клюва 0,9-1,2 см. Длина
крыла самок 6-7,5 см, хвоста 5-6 см, цевки 1,5-2 см, клюва 0,9-1,2 см.
В июне – июле воробей пробуждается за 30–40 мин. до восхода солнца. В течение светлой части суток периоды двигательной (кормовой) активности сменяются периодами относительного покоя. Такого рода активность выше в утренние (8–11 ч.) и вечерние (18–20 ч.) часы, в остальное время суток она несколько ниже.
Населяет культурный ландшафт - деревни, усадьбы, а также города (не только окраины, но и центральные части, однако здесь менее обычен). Кроме того, довольно много полевых воробьев гнездится в
поймах рек, разреженных лесах, колониальных поселениях грачей и серых цапель. В гнездовой период придерживается парков, садов, древесных насаждений среди полей, пойм рек, опушек старых лесов
(главным образом лиственных), строений сельского типа. Гнездится часто по соседству или недалеко от селений человека.
Репродуктивный период полевого воробья состоит из 2-3 циклов размножения. Каждый из циклов включает 6 последовательных фаз: токование, строительство гнезда, откладка яиц, насиживание, выкармливание птенцов в гнезде и вождение выводка. Во втором и третьем циклах размножения такие фазы, как токование и гнездостроение, зачастую значительно сокращаются или полностью выпадают.
Начало весеннего токования у полевого воробья сильно варьирует во времени (первая – третья декады марта) и зависит, прежде всего, от фотопериода и температуры.
Время от начала токования до начала постройки гнезда у полевого воробья составляет от 10 до 25 дней. Сроки начала гнездования варьируют по годам в пределах одного месяца.
Продолжительность гнездостроения у воробьев изменяется от 5 до 20 и более дней, в большинстве случаев составляет 7–15 дней. Птицы перед началом второй кладки ремонтируют гнезда, ремонт
продолжается около 4–8 дней. Воробьи часто (около 20%) бросают недостроенные гнезда и возводят гнездо в другом месте.
Полевые воробьи гнездятся колониями. Гнезда устраивают в дуплах деревьев, в искусственных гнездовьях (скворечники, синичники), в щелях и под крышами домов, за наличниками, в гнездах белого аиста, хищников, ласточек, изредка в береговых обрывах, занимая пустоты, ниши и норы, вырытые береговыми ласточками. Гнездо, как правило, объемистое, рыхлое. Размеры его зависят от величины полости, в которой оно помещается. Строительным материалом служат сухие стебли растений (нередко полуистлевшие), солома, корешки, летучие и пушистые семена и т. д. Выстилка очень обильная, состоит из большого количества перьев, пуха, шерсти и иного мягкого материала. Высота гнезда 5–8 см, диаметр 11–14 см, глубина лотка 3–6 см, диаметр лотка 4,5–6 см.
В полной кладке 4-6 (чаще 4-5), иногда 7 яиц. Размеры, форма и окраска их сильно варьируют. Скорлупа матовая либо слегка блестящая. На белом, голубоватом или зеленовато-белом фоне
густо рассеяны различной формы и величины пятнышки. Они обычно бывают оливкового, светло- и зеленовато-бурого, реже коричневого цвета. Вес яйца 2,1 г, длина 16-22 мм, диаметр 12-15 мм.
Вес яйца 2,1 г, длина 16-22 мм, диаметр 12-15 мм.
К откладке яиц птица приступает в первой декаде мая. Воробьи даже в одной колонии приступают к откладке яиц в разные сроки, этот процесс растягивается иногда на 15–20 суток и более. В году у некоторых пар бывает до трех выводков. Свежие кладки встречаются до середины июля. Насиживают кладку в течение 11-14 суток обе птицы, столько же времени птенцы находятся в гнезде (по другим данным, 15–16 суток). Родители выкармливают птенцов мелкими животными и растительными кормами. В рационе птенцов преобладают жесткокрылые, гусеницы чешуекрылых и тли. Количество беспозвоночных, идущее на выкармливание выводка из 5 птенцов, составляет 5,7–8,2 тыс. экз.
В первые дни жизни птенцы получают корм редко, но по мере их роста интенсивность кормления возрастает. В возрасте птенцов 3–4 дня количество прилетов родителей к гнезду с кормом составляет
166–170 раз в день, в возрасте 8–10 дней – 190–220 раз, за 2–3 дня перед вылетом – 250–290 раз. Интенсивность кормления птенцов изменяется в течение светлой части суток. Наибольшая она в утренние
и послеобеденные (вечерние) часы, несколько снижается в середине дня. В дождливые, пасмурные и ветреные дни интенсивность кормления снижается в 1,5–2 раза.
Интенсивность кормления птенцов изменяется в течение светлой части суток. Наибольшая она в утренние
и послеобеденные (вечерние) часы, несколько снижается в середине дня. В дождливые, пасмурные и ветреные дни интенсивность кормления снижается в 1,5–2 раза.
Интенсивность прилетов родителей с кормом зависит от количества птенцов в гнезде. К гнездам, в которых находится 5 и более птенцов, взрослые птицы с кормом прилетают значительно чаще по сравнению с гнездами, количество птенцов в которых составляет 2–4.
После вылета птенцов полевые воробьи концентрируются в стаи, иногда довольно большие (до 200 особей), летают кормиться на поля и свалки.
Пищевой спектр этого вида примерно тот же, что и у домового воробья. Но в летнее время животные корма (в первую очередь насекомые) играют, пожалуй, более значимую роль. В частности, птенцы в
гнездах выкармливаются почти исключительно насекомыми и пауками. В питании взрослых птиц вне периода размножения преобладают семена культурных и дикорастущих растений.
Кормовая активность птиц в весенне-летний период выше в утренние и вечерние часы. Зимой воробьи кормятся на протяжении всего светлого времени суток. Пик кормовой активности приходится на середину дня. Часто птицы добывают корм в сотрудничестве с врановыми (грачи, галки, серые вороны), которые способны раскапывать клювом снег. Полевые воробьи могут сами добывать корм из-под снега, высота которого не превышает 10 см.
В течение года численность птиц наибольшая летом и осенью, наименьшая – зимой и весной. Во внегнездовой период птицы совершают кочевки в поисках корма. Осенью и зимой наибольшая плотность
населения птиц наблюдается в сельских населенных пунктах, на полях зерновых, где они находят семена культурных растений, а также на полях, заросших сорняками. Поздней осенью и в начале зимы, с
установлением снегового покрова стаи полевых воробьев распадаются на небольшие группы (15–30 особей) и концентрируются у животноводческих ферм или в населенных пунктах, где находят корма
антропогенного происхождения. При этом птицы придерживаются окраин поселений, примыкающих к полям, пустырям. В центре городов концентрируются около рынков и магазинов. Видимо, значительная часть
популяции не ограничивается местными кочевками, а мигрирует на зиму в более южные районы, возвращается к местам гнездовий весной.
При этом птицы придерживаются окраин поселений, примыкающих к полям, пустырям. В центре городов концентрируются около рынков и магазинов. Видимо, значительная часть
популяции не ограничивается местными кочевками, а мигрирует на зиму в более южные районы, возвращается к местам гнездовий весной.
Численность полевого воробья в последние десятилетия в Беларуси оценивается в 900–950 тыс. гнездящихся пар. За последние 22 года она несколько раз снижалась. Отмечено слабое увеличение численности полевого воробья в последние 5 лет.
Максимальный зарегистрированный в Европе возраст 13 лет 1 месяц.
Gurkov2N. Витебский р-н
Все, что вам нужно знать о SPARROW
Посетите национальный веб-сайт USGS SPARROW, чтобы ознакомиться с обзором моделирования SPARROW.
ВОРОБЕЙ Техника моделирования
Ученые Геологической службы США разработали SPARROW (Смит и др., 1997 г.) до:
- Использование данных мониторинга и информации о водоразделах для лучшего объяснения факторов, влияющих на качество воды
- Изучение статистической значимости источников загрязнителей, факторов окружающей среды и процессов переноса для объяснения прогнозируемых нагрузок загрязняющих веществ
- Обеспечить статистическую основу для оценки водотоков в неконтролируемых местах

Модель оценивает концентрации загрязняющих веществ, потоки (или «массу», которая является произведением концентрации и речного стока) и выходы в ручьях (масса питательных веществ, поступающих в ручей на акр земли), а также оценивает вклад выбранных источников загрязняющих веществ и свойства водосборных бассейнов, которые контролируют транспорт в крупных речных сетях. Он эмпирически оценивает происхождение и судьбу загрязняющих веществ в ручьях и принимающих телах и количественно определяет неопределенности в этих оценках на основе погрешности коэффициента и необъяснимой изменчивости наблюдаемых данных.
Модель SPARROW основана на фактическом мониторинге ручья с использованием всеобъемлющих геопространственных данных в откалиброванной модели SPARROW для прогнозирования условий качества воды в неконтролируемых местах ручья (см. иллюстрацию ниже). Наборы геопространственных данных описывают применение удобрений и навоза, атмосферное осаждение на поверхность земли и городские источники. Прогнозы модели иллюстрируются подробными картами, которые предоставляют информацию о нагрузках загрязняющих веществ и вкладе источников в различных масштабах для конкретных участков водотоков, бассейнов или других географических областей.
Прогнозы модели иллюстрируются подробными картами, которые предоставляют информацию о нагрузках загрязняющих веществ и вкладе источников в различных масштабах для конкретных участков водотоков, бассейнов или других географических областей.
SPARROW и отдельные результаты для водосборных бассейнов США представлены Смитом и другими, 1997. Теорию, модельную документацию и иллюстрированное пользовательское приложение SPARROW можно найти в онлайн-отчете о методах USGS, The SPARROW Surface Water-Quality Model. : Теория, применение и пользовательская документация от GE Шварц, А.Б. Хус, Р.Б. Александер и Р.А. Смит.
Как измерения нагрузки потока используются в SPARROW?
Информация о загрузке потока используется для калибровки моделей SPARROW и, таким образом, является для них одним из наиболее важных типов данных.Чтобы максимизировать точность и точность калибровки модели SPARROW, важно иметь набор измерений нагрузки, которые являются репрезентативными для рассматриваемой области и доступны в достаточном количестве, чтобы зафиксировать изменчивость, которая происходит в этой области. По этим причинам значительные усилия затрачиваются на поиск загружаемых данных со всех смоделированных водоразделов (и даже близлежащих водосборов). Эти усилия часто требуют интеграции данных из различных источников, включая федеральные, государственные и местные агентства по управлению качеством воды.
По этим причинам значительные усилия затрачиваются на поиск загружаемых данных со всех смоделированных водоразделов (и даже близлежащих водосборов). Эти усилия часто требуют интеграции данных из различных источников, включая федеральные, государственные и местные агентства по управлению качеством воды.
Для получения информации о нагрузке на реку собираются данные о качестве воды по как можно большему количеству участков в интересующей области. Информация о сбросе воды с гидрометрических постов также собирается, а расположение водомеров сопоставляется с пунктами мониторинга качества воды, чтобы данные о сбросе можно было связать с данными о концентрации воды. Статистические процедуры используются для оценки годовых нагрузок на основе соотношения сток-концентрация и временных изменений, таких как тренды (Schwarzand others, 2006).В большинстве случаев используются долгосрочные данные о расходе воды, а оценки нагрузки «детрендированы» для оценки нагрузки на реку, которая могла бы возникнуть при текущих условиях качества воды, обусловленных долгосрочными средними условиями расхода. Эта процедура удаления тренда устраняет пространственные вариации, которые могут быть связаны с погодными условиями, и помогает сфокусировать модель исключительно на пространственных закономерностях факторов окружающей среды, влияющих на качество воды.
Эта процедура удаления тренда устраняет пространственные вариации, которые могут быть связаны с погодными условиями, и помогает сфокусировать модель исключительно на пространственных закономерностях факторов окружающей среды, влияющих на качество воды.
Какие типы данных используются для прогнозирования в моделях SPARROW?
Моделирование SPARROW требует интеграции многих типов геопространственных данных для использования в качестве независимых переменных, которые рассматриваются либо как составные источники, либо как факторы доставки.Источники могут включать определенные типы земель, такие как городские районы, или известные источники загрязнения, такие как очистные сооружения. Условия поставки могут включать любые характеристики бассейна, которые могут быть связаны с естественным затуханием. Например, денитрификация часто связана с определенными характеристиками почвы, а пространственная структура этих характеристик почвы часто связана с составляющими нагрузками. В некоторых случаях условия доставки также могут быть связаны с расширенной доставкой. Например, высокий уклон бассейна может привести к более быстрому течению, что может увеличить поступление компонентов.На доставку также влияет время прохождения по воде в ручьях, которое можно оценить по опубликованным исследованиям времени прохождения Геологической службы США (например, Reed and Stuckey, 2001). Примеры некоторых наборов геопространственных данных, использованных для разработки независимых переменных в прошлых моделях SPARROW, перечислены ниже.
В некоторых случаях условия доставки также могут быть связаны с расширенной доставкой. Например, высокий уклон бассейна может привести к более быстрому течению, что может увеличить поступление компонентов.На доставку также влияет время прохождения по воде в ручьях, которое можно оценить по опубликованным исследованиям времени прохождения Геологической службы США (например, Reed and Stuckey, 2001). Примеры некоторых наборов геопространственных данных, использованных для разработки независимых переменных в прошлых моделях SPARROW, перечислены ниже.
Наборы данных об источнике загрязнения для:
Наборы данных о доставке загрязняющих веществ:
Какие источники качества воды учитываются в моделях SPRROW?
Многие факторы окружающей среды были идентифицированы как источники в моделях SPARROW.К ним относятся точечные источники, определяемые наборами данных, описывающими фактические местоположения сбросов, таких как очистные сооружения, или наборами данных, описывающими население или городскую территорию в качестве заменителей точечных источников. Сельскохозяйственные источники также были идентифицированы в различных формах, включая:
Сельскохозяйственные источники также были идентифицированы в различных формах, включая:
- Сельскохозяйственные земли, определенные с использованием данных о землепользовании/почвенном покрове
- Оценка применения удобрений
- Оценки образования навоза
- Оценки питательных веществ, применяемых к конкретным культурам.
Другие источники, выявленные с помощью моделей SPARROW, включают атмосферные выпадения, городские земли и естественные источники, такие как районы добычи полезных ископаемых. Во всех случаях эти наборы данных постоянно улучшаются, и эти улучшения включаются в модели SPARROW по мере их появления.
Любой из этих составных источников или других может быть потенциально включен в модель SPARROW при условии, что доступны геопространственные данные для его описания и пространственные закономерности в источнике могут быть успешно сопоставлены с характеристиками в измерениях потоковой нагрузки этого составляющего.
Поскольку СПАРРОУ основан на балансе масс, источники должны быть доступны для всех частей региона, чтобы определить их общую важность. Таким образом, некоторые наборы данных, которые предоставляют подробную информацию только для части области модели, не будут полезны в модели SPARROW, поскольку одна и та же информация не будет доступна везде. Например, подробные оценки сельскохозяйственных затрат азота, собранных в одном штате, могут оказаться бесполезными для модели, охватывающей всю страну, поскольку в других штатах данные будут отсутствовать.
Таким образом, некоторые наборы данных, которые предоставляют подробную информацию только для части области модели, не будут полезны в модели SPARROW, поскольку одна и та же информация не будет доступна везде. Например, подробные оценки сельскохозяйственных затрат азота, собранных в одном штате, могут оказаться бесполезными для модели, охватывающей всю страну, поскольку в других штатах данные будут отсутствовать.
Успешная корреляция источника с измерениями нагрузки потока (и его включение в модель SPARROW в качестве статистически значимого источника) зависит от того,
Источник достаточно велик, чтобы вносить важный вклад в общий баланс массы в сети водотоков, и (b) пространственная изменчивость в этом источнике, описываемая наборами геопространственных данных, достаточно велика. Оба эти условия необходимы для того, чтобы SPRROW идентифицировал источник как статистически значимый, т.е.е., обнаружить, что пространственные закономерности в источнике коррелируют с таковыми в качестве воды в реке.
Как характеристики водосбора используются в моделях SPARROW?
Таким же образом, как SPARROW определяет относительную важность источников загрязнения для рек, он также оценивает важность факторов ландшафта в доставке этих загрязнителей в ручьи. SPARROW налагает ограничения баланса массы на все оценки загрязнения загрязняющими веществами. Таким образом, все источники должны быть уравновешены процессами затухания окружающей среды (потери), чтобы оценить измерения нагрузки потока с минимальной ошибкой.Например, питательные вещества, происходящие из сельскохозяйственных угодий, могут быть потеряны в результате денитрификации, поскольку они переносятся с поверхности земли через неглубокие грунтовые воды в ручьи. Проницаемость почвы может усиливать денитрификацию, поэтому пространственное распределение значения проницаемости почвы может быть связано с нагрузкой загрязнителей ниже по течению.
Любая характеристика ландшафта может быть оценена как потенциальный фактор потерь при попадании загрязняющих веществ в ручьи. Ландшафтные характеристики могут быть статистически идентифицированы как важные только в том случае, если они достаточно различаются по моделируемой территории и их можно отличить по своей величине от пространственной изменчивости других ландшафтных характеристик, которые не менее важны.Например, проницаемость почвы и органическое вещество почвы не были бы идентифицированы как статистически важные, если бы они были связаны с одним и тем же основным процессом затухания (например, денитрификацией) и происходили в одной и той же пространственной структуре.
Ландшафтные характеристики могут быть статистически идентифицированы как важные только в том случае, если они достаточно различаются по моделируемой территории и их можно отличить по своей величине от пространственной изменчивости других ландшафтных характеристик, которые не менее важны.Например, проницаемость почвы и органическое вещество почвы не были бы идентифицированы как статистически важные, если бы они были связаны с одним и тем же основным процессом затухания (например, денитрификацией) и происходили в одной и той же пространственной структуре.
Многие характеристики ландшафта были идентифицированы как важные факторы затухания в моделях SPARROW. Примеры включают характеристики почв, такие как проницаемость почвы, климатические факторы, такие как долгосрочная средняя температура и осадки, физико-географические характеристики, такие как уклон и топография, и схемы дренажа, такие как плотность потока и искусственный дренаж.Новые наборы данных продолжают разрабатываться и, таким образом, создают возможности для оценки новых потенциальных факторов потерь в моделях SPARROW.
Какова пространственная структура, лежащая в основе моделей SPARROW?
SPARROW предназначен для описания пространственных закономерностей качества воды и факторов, влияющих на него. Это достигается путем связывания участков мониторинга качества воды с цифровой сетью водотоков, которая описывает пространственные связи между участками (вверх или вниз по течению).Сети цифровых потоков предоставляют информацию о потоках по всему региону и обычно разбивают потоки на сегменты (досягаемости), которые различаются по размеру в зависимости от масштаба набора данных. Границы водосбора устанавливаются для каждого участка ручья, и эти границы используются для определения участков водосбора, вносящих вклад в участки ручья. Базы географических данных, описывающие характеристики водосборных бассейнов, такие как землепользование, связаны с водосборными площадями для количественной оценки количества каждого фактора, влияющего на условия на каждом участке реки. Таким образом, охват цифрового водотока и связанные с ним водосборные площади обеспечивают пространственную структуру, которая позволяет интегрировать данные, собранные на участках мониторинга качества воды, с данными, описывающими характеристики водосборных бассейнов вверх по течению.
Таким образом, охват цифрового водотока и связанные с ним водосборные площади обеспечивают пространственную структуру, которая позволяет интегрировать данные, собранные на участках мониторинга качества воды, с данными, описывающими характеристики водосборных бассейнов вверх по течению.
Stream Reach Networks : На сегодняшний день в моделях SPARROW преимущественно использовались две цифровые потоковые сети (см. рисунок ниже). Первая называется У.S. Enhanced Reach File 1 или ERF1 Агентства по охране окружающей среды (Александр и другие, 1999). Этот набор данных распространяется на континентальную часть Соединенных Штатов и включает приблизительно 62 000 рек в масштабе 1:500K. Вторая сеть цифровых водотоков, используемая в моделях SPARROW, известна как Национальный набор гидрографических данных (NHD) (Геологическая служба США, 1999 г.). Однако теперь доступна более новая расширенная версия, известная как NHDPlus, которая включает многие атрибуты охвата потока, которые не были доступны в исходном наборе данных NHD. Как и ERF1, NHD (и NHDPlus) распространяется на континентальную часть Соединенных Штатов. Однако NHD был разработан для включения гораздо большего количества пространственных деталей и включает 2,6 миллиона потоков в масштабе 1:100K.
Как и ERF1, NHD (и NHDPlus) распространяется на континентальную часть Соединенных Штатов. Однако NHD был разработан для включения гораздо большего количества пространственных деталей и включает 2,6 миллиона потоков в масштабе 1:100K.
Как характеристики ландшафта и данные мониторинга связаны с пространственной структурой в моделях SPARROW?
SPARROW предоставляет инструмент прогнозирования, который объединяет множество типов данных. Калибровочные данные получены на основе информации о мониторинге качества воды на участках, расположенных по всей изучаемой территории.Эти данные связаны с участками в сети цифровых водотоков для определения пространственных отношений между участками мониторинга и между их водосборными площадями. Затем подробные базы геопространственных данных связываются с водосборными площадями речной сети для определения характеристик бассейнов во всех областях, впадающих в места мониторинга и на все участки отдельных ручьев. После установления связей все эти типы данных объединяются в одну базу данных, которая используется для разработки модели.
После установления связей все эти типы данных объединяются в одну базу данных, которая используется для разработки модели.
На рисунке ниже представлен небольшой пример и иллюстрация пространственной структуры, которая составляет основу SPARROW, как описано выше.
Темно-черная линия представляет собой границу дренажа водораздела. В нижней части водораздела находится место мониторинга, которое можно использовать для калибровки модели. Выше по течению от места мониторинга водотоки в водоразделе разбиты на ряд участков, называемых участками ручьев. Границы водосбора и области, вносящие вклад в участки ручья, определены и показаны отдельными цветами. В пределах каждой области экологические факторы, влияющие на качество воды, количественно оцениваются с использованием пространственно детализированных географических данных.Факторы окружающей среды могут включать в себя широкий спектр источников, таких как очистные сооружения, городские районы или сельскохозяйственные угодья. Факторы окружающей среды также могут включать характеристики, связанные с естественным ослаблением (утратой) загрязняющих веществ посредством таких процессов, как денитрификация или секвестрация. Примеры таких факторов окружающей среды включают проницаемость почвы или геологические характеристики. Второй тип потерь, учитываемых в SPARROW, — это потери, возникающие при прохождении через потоковые каналы.Потери на входе оцениваются путем сравнения измерений нагрузки на входе и на выходе. Нагрузки загрязняющих веществ, поступающие из верхнего течения, а также из промежуточных участков ручья, уравновешиваются измеренной нагрузкой вниз по течению для оценки величины потерь в русле. Таким образом, массовый баланс обеспечивает фундаментальную основу SPARROW.
Факторы окружающей среды также могут включать характеристики, связанные с естественным ослаблением (утратой) загрязняющих веществ посредством таких процессов, как денитрификация или секвестрация. Примеры таких факторов окружающей среды включают проницаемость почвы или геологические характеристики. Второй тип потерь, учитываемых в SPARROW, — это потери, возникающие при прохождении через потоковые каналы.Потери на входе оцениваются путем сравнения измерений нагрузки на входе и на выходе. Нагрузки загрязняющих веществ, поступающие из верхнего течения, а также из промежуточных участков ручья, уравновешиваются измеренной нагрузкой вниз по течению для оценки величины потерь в русле. Таким образом, массовый баланс обеспечивает фундаментальную основу SPARROW.
Что такое базовый год и почему он используется в моделях SPARROW?
В целом, прогнозы источников питательных веществ и нагрузок модели SPARROW отражают долгосрочные среднегодовые условия содержания питательных веществ в ручьях. Статистическая процедура используется для обеспечения того, чтобы прогнозы модели отражали долгосрочную гидрологическую изменчивость и изменчивость качества воды в течение согласованного периода времени, что дает надежные прогнозы модели источников питательных веществ и процессов переноса. Модельные прогнозы среднегодовой нагрузки для откалиброванной модели стандартизированы для одного года, называемого «базовым годом», чтобы дать оценку средней нагрузки биогенными веществами, которая имела бы место в реках в течение этого года, если бы среднегодовые условия стока были победил.
Статистическая процедура используется для обеспечения того, чтобы прогнозы модели отражали долгосрочную гидрологическую изменчивость и изменчивость качества воды в течение согласованного периода времени, что дает надежные прогнозы модели источников питательных веществ и процессов переноса. Модельные прогнозы среднегодовой нагрузки для откалиброванной модели стандартизированы для одного года, называемого «базовым годом», чтобы дать оценку средней нагрузки биогенными веществами, которая имела бы место в реках в течение этого года, если бы среднегодовые условия стока были победил.
Назначенный базовый год для модели SPARROW обычно выбирается для обеспечения согласованности с другими вспомогательными данными, используемыми в модели, включая данные об источниках питательных веществ, землепользовании, климате, сети водотоков и т. д. Данные о земном покрове (Национальные данные о земном покрове Геологической службы США). или NLCD) доступны только за определенные годы, обычно с интервалом в 5 лет. Они дополняются сельскохозяйственными статистическими данными о питательных веществах для животных, посевных площадях и производстве, которые публикуются только каждые 5 лет. Оценки содержания питательных веществ в навозе животных, основанные на округах, из U.Министерство сельского хозяйства США (USDA) также сообщается только за определенные периодические годы.
Они дополняются сельскохозяйственными статистическими данными о питательных веществах для животных, посевных площадях и производстве, которые публикуются только каждые 5 лет. Оценки содержания питательных веществ в навозе животных, основанные на округах, из U.Министерство сельского хозяйства США (USDA) также сообщается только за определенные периодические годы.
Каким образом долгосрочные данные о стоке и качестве воды интегрируются для расчета годовой нагрузки на реку?
В модели SPARROW для калибровки модели используется среднегодовая нагрузка (масса в единицу времени, выраженная, например, в килограммах в год) на каждой станции мониторинга водотоков. Среднегодовая нагрузка рассчитывается с использованием методов оценки нагрузки Геологической службы США; подробности см. в документации модели SPARROW.
Вкратце, эти методы оценки нагрузки сочетают в себе регулярно собираемые значения биогенных веществ на станциях мониторинга с ежедневными значениями речного стока, как правило, за предыдущий 30-летний период. Этот подход дает более точные оценки долгосрочной среднегодовой нагрузки, чем те, которые можно получить, используя только измерения отдельных загрязняющих веществ. Среднегодовая нагрузка загрязняющими веществами также стандартизирована по единому времени, называемому «базовым годом», который является назначенным периодом времени для сравнения одновременной пространственной изменчивости нагрузок на качество воды и источников загрязнения в модели SPARROW.Стандартизация обеспечивает оценку средней нагрузки загрязняющими веществами, которая имела бы место в установленном базовом году, если бы преобладали условия среднегодового стока. Использование стандартизированных нагрузок в модели SPARROW дает более надежные оценки источников загрязнения и процессов переноса по сравнению с подходом, основанным исключительно на использовании записей о качестве воды в течение любого одного года или коротких многолетних периодов. Процедура стандартизации учитывает различия в длинах записей станций и размерах выборок и гарантирует, что биогенные нагрузки репрезентативны для долгосрочной гидрологической изменчивости и изменчивости качества воды.
Этот подход дает более точные оценки долгосрочной среднегодовой нагрузки, чем те, которые можно получить, используя только измерения отдельных загрязняющих веществ. Среднегодовая нагрузка загрязняющими веществами также стандартизирована по единому времени, называемому «базовым годом», который является назначенным периодом времени для сравнения одновременной пространственной изменчивости нагрузок на качество воды и источников загрязнения в модели SPARROW.Стандартизация обеспечивает оценку средней нагрузки загрязняющими веществами, которая имела бы место в установленном базовом году, если бы преобладали условия среднегодового стока. Использование стандартизированных нагрузок в модели SPARROW дает более надежные оценки источников загрязнения и процессов переноса по сравнению с подходом, основанным исключительно на использовании записей о качестве воды в течение любого одного года или коротких многолетних периодов. Процедура стандартизации учитывает различия в длинах записей станций и размерах выборок и гарантирует, что биогенные нагрузки репрезентативны для долгосрочной гидрологической изменчивости и изменчивости качества воды. Этот акцент на долгосрочных средних условиях расширяет возможности модели для оценки основных источников и водосборных процессов, которые влияют на долгосрочное снабжение, перенос и судьбу питательных веществ в водосборных бассейнах.
Этот акцент на долгосрочных средних условиях расширяет возможности модели для оценки основных источников и водосборных процессов, которые влияют на долгосрочное снабжение, перенос и судьбу питательных веществ в водосборных бассейнах.
Как учитываются временные вариации количества осадков и гидрология в моделях SPARROW?
Фактические измерения нагрузки на качество воды показывают большие ежегодные колебания, в основном обусловленные ежегодными изменениями погодных условий и условий стока.Например, долгосрочный мониторинг качества воды и речного стока Геологической службы США показывает, что количество азота увеличилось с конца 1960-х годов; и что это количество было низким во время засухи в конце 1980-х годов, но высоким во время наводнения 1993 года, даже несмотря на то, что количество азота, внесенного на поля в бассейне, существенно не отличалось (см. Информационный бюллетень 135-00). Кроме того, межгодовые вариации среднегодовой нагрузки азотом могут достигать двух порядков на станциях мониторинга, хотя годовые вариации обычно составляют 20-40 процентов от многолетней среднегодовой нагрузки. .
.
Годовые вариации, подобные этим, и внутригодовые вариации концентрации загрязняющих веществ и речного стока учитываются на этапе, предшествующем пространственному моделированию SPARROW. Этот предварительный шаг имеет решающее значение для получения нагрузок загрязняющих веществ в реке для калибровки модели SPARROW, которые представляют долгосрочные средние гидрологические условия и качество воды на каждой станции мониторинга. SPARROW имеет целью объяснить пространственные различия в нагрузках загрязняющих веществ, которые связаны с внутренними факторами, которые контролируют средние скорости поступления и переноса питательных веществ, а не с факторами, которые объясняют более экстремальные гидрологические условия в течение какого-либо конкретного года или лет в определенном месте.Прогнозы SPARROW по средней нагрузке питательными веществами включают влияние пространственных вариаций средних климатических условий (осадки, температура) и речного стока. Этот акцент на долгосрочных средних условиях расширяет возможности пространственной модели для оценки основных источников загрязнения, включая землепользование и деятельность человека, а также естественные процессы, влияющие на долгосрочное снабжение, перенос и судьбу питательных веществ в водосборных бассейнах.
Можно ли использовать модель SPARROW для оценки тенденций в нагрузках загрязняющих веществ в ручьях?
Предыдущие модели SPARROW были разработаны для описания среднегодовых нагрузок на качество воды для определенного базового года и не использовались для описания тенденций или изменений во времени нагрузок на водотоки.Тем не менее, текущие исследования сосредоточены на разработке моделей SPARROW десятилетних и сезонных изменений нагрузки на реку. Модели десятилетних изменений будут предполагать, что источники, процессы и водотоки находятся в устойчивом состоянии в течение каждого моделируемого 10-летнего периода. Таким образом, эти модели дают возможность оценить, какая часть изменения нагрузки рек является результатом изменений поступления загрязняющих веществ из различных источников (например, сбросов очистки сточных вод, внесения удобрений) по сравнению с той частью, которая возникает в результате основных изменений в процессах и деятельности человека, которые может изменить скорость поступления загрязняющих веществ в водотоки (т. г., агроприемы консервации, минерализация питательных веществ в почве).
г., агроприемы консервации, минерализация питательных веществ в почве).
ВОРОБЕЙ Моделирование: предсказание и неопределенность
Какие показатели предсказывают модели SPARROW?
МоделиSPARROW разрабатываются с использованием ограничений баланса массы для количественной оценки взаимосвязи между составной нагрузкой потока (массой составляющей, переносимой потоком) и источниками и потерями массы в водоразделах. Таким образом, модели по своей сути предназначены для прогнозирования нагрузки (массы за время) для всех участков потока в моделируемом регионе.Тем не менее, прогнозы нагрузки на реку могут быть изменены, чтобы обеспечить различные показатели качества воды, которые могут поддерживать различные типы оценок.
Показатели прогнозирования SPARROW включают составляющую урожайность, концентрацию и вклад источника в нагрузки потока:
(a) Составляющие выходы:
Урожайность составляющих (масса на единицу площади за время) рассчитывается как речная нагрузка, деленная на площадь водосбора. Меры урожайности полезны для менеджеров по качеству воды для определения относительного вклада загрязняющих веществ из разных частей большой площади водосбора.Нагрузка загрязняющих веществ обычно увеличивается с площадью водосбора ручья, что затрудняет сравнение нагрузок между ручьями разного размера. Урожайность дает меру речной нагрузки, нормализованную по размеру дренажа, и, таким образом, обеспечивает надежную метрику для определения того, какие дренажи выносят наибольшее количество загрязняющих веществ независимо от их размера.
Меры урожайности полезны для менеджеров по качеству воды для определения относительного вклада загрязняющих веществ из разных частей большой площади водосбора.Нагрузка загрязняющих веществ обычно увеличивается с площадью водосбора ручья, что затрудняет сравнение нагрузок между ручьями разного размера. Урожайность дает меру речной нагрузки, нормализованную по размеру дренажа, и, таким образом, обеспечивает надежную метрику для определения того, какие дренажи выносят наибольшее количество загрязняющих веществ независимо от их размера.
SPARROW позволяют прогнозировать «общую», «дополнительную» и «доставленную» урожайность на основе соответствующих показателей нагрузки. Эти меры предоставляют важную для управления информацию об источниках и судьбе загрязняющих веществ в различных пространственных масштабах.«Общая» мощность описывает нагрузку на единицу площади, которая доставляется к нижнему концу участка от источников по всему дренажу над участком (т. е. рассчитывается как общая нагрузка, доставляемая до конца участка, деленная на общую площадь водосбора вверх по течению, связанная с досягаемостью). Напротив, «дополнительный» сток описывает нагрузку на единицу площади, доставляемую к нижнему концу участка исключительно из источников на земельном участке, который стекает непосредственно в участок, не проходя через другой участок (т.д., «дополнительный» дренаж). Таким образом, дополнительный выход не зависит от вклада загрязняющих веществ из водосборных площадей верхних участков и измеряет относительно «местные» источники загрязняющих веществ, которые попадают в один участок ручья.
Напротив, «дополнительный» сток описывает нагрузку на единицу площади, доставляемую к нижнему концу участка исключительно из источников на земельном участке, который стекает непосредственно в участок, не проходя через другой участок (т.д., «дополнительный» дренаж). Таким образом, дополнительный выход не зависит от вклада загрязняющих веществ из водосборных площадей верхних участков и измеряет относительно «местные» источники загрязняющих веществ, которые попадают в один участок ручья.
Наконец, «доставленный» выход полезен для описания судьбы загрязняющей нагрузки на единицу площади ниже по течению для данного участка реки. Этот показатель особенно полезен для количественной оценки вклада водосборных бассейнов в загрязняющую нагрузку (на основе общей или дополнительной площади водосбора) в принимающие воды ниже по течению, такие как водохранилища или чувствительные прибрежные эстуарии.Доставленный выход учитывает естественное ослабление загрязняющих веществ в ручьях и водоемах (например, длительное хранение; денитрификация) во время транспортировки от данного участка ручья до принимающих вод ниже по течению. Доставленный сток рассчитывается путем умножения «общего» или «добавочного» стока ручья на оценку «доли доставки» по модели SPARROW, которая описывает долю загрязняющей нагрузки, которая не ослабляется или не удаляется естественными процессами во время нисходящий транспорт.Существует много примеров использования данных об урожае SPARROW для определения водосборных бассейнов, оказывающих наибольшую нагрузку на прибрежные воды, в том числе бассейнов рек Миссисипи, Чесапикского залива и водосборных бассейнов Новой Англии.
Доставленный сток рассчитывается путем умножения «общего» или «добавочного» стока ручья на оценку «доли доставки» по модели SPARROW, которая описывает долю загрязняющей нагрузки, которая не ослабляется или не удаляется естественными процессами во время нисходящий транспорт.Существует много примеров использования данных об урожае SPARROW для определения водосборных бассейнов, оказывающих наибольшую нагрузку на прибрежные воды, в том числе бассейнов рек Миссисипи, Чесапикского залива и водосборных бассейнов Новой Англии.
(b) Концентрации в составе:
Концентрация определяется в SPARROW путем деления прогнозируемой нагрузки на долгосрочный средний расход в каждом участке ручья. Меры концентрации наиболее полезны для понимания пригодности воды для использования водными организмами и людьми (например,g., контакт с водой, снабжение питьевой водой) и в решающей степени зависят от объема воды, протекающей в ручьях. Важно отметить, что прогнозы концентрации SPARROW являются взвешенными по потоку оценками. Эти оценки могут отличаться от взвешенных по времени оценок концентраций качества воды, которые АООС США и штаты часто используют для оценки соблюдения нормативных стандартов (дополнительная информация доступна в исследованиях USGS взвешенных по времени концентраций; например, Эффекты обогащения питательными веществами в ручьях; Концентрация питательных веществ в ручьях; Тенденции в обогащении ручьев питательными веществами).
Эти оценки могут отличаться от взвешенных по времени оценок концентраций качества воды, которые АООС США и штаты часто используют для оценки соблюдения нормативных стандартов (дополнительная информация доступна в исследованиях USGS взвешенных по времени концентраций; например, Эффекты обогащения питательными веществами в ручьях; Концентрация питательных веществ в ручьях; Тенденции в обогащении ручьев питательными веществами).
(c) Вклад источников в речные нагрузки:
Специалистам по качеству воды также необходимо определять вклад загрязняющих веществ в водотоки, происходящие из различных типов источников в водосборных бассейнах. SPARROW предоставляет оценки основных источников загрязняющих веществ в ручьях, включая муниципальные точечные источники, а также городские и сельскохозяйственные диффузные источники. Предсказываемые моделью вклады источников в потоки могут быть выражены в виде нагрузки, выхода или в относительном выражении в процентах от нагрузки потока. Кроме того, вклад источника может быть указан для «общей» или «добавочной» площади водосбора, связанной с участками ручья. В совокупности этот высокоинформативный набор модельных прогнозов можно использовать для оценки «местного» и регионального вклада основных источников загрязнения как во внутренние водотоки, так и в прибрежные воды.
Кроме того, вклад источника может быть указан для «общей» или «добавочной» площади водосбора, связанной с участками ручья. В совокупности этот высокоинформативный набор модельных прогнозов можно использовать для оценки «местного» и регионального вклада основных источников загрязнения как во внутренние водотоки, так и в прибрежные воды.
Как пространственный масштаб влияет на предсказания модели SPARROW?
Модели SPARROW, как правило, разрабатываются как «независимые от масштаба», что означает, что прогнозы модели считаются действительными в широком диапазоне пространственных масштабов, приблизительно соответствующих масштабу геопространственных данных, используемых для калибровки модели.Как правило, модели можно использовать для моделирования условий качества воды в масштабах, варьирующихся от водосборных бассейнов одного ручья (от нескольких квадратных километров до десятков квадратных километров) до размеров крупных водоразделов и речных бассейнов, охватывающих тысячи квадратных километров. . Таким образом, модели SPARROW являются гибкими и могут использоваться для выполнения различных типов оценок, охватывающих диапазон размеров бассейнов.
. Таким образом, модели SPARROW являются гибкими и могут использоваться для выполнения различных типов оценок, охватывающих диапазон размеров бассейнов.
Однако существуют связанные с масштабом ограничения на использование модели для управления качеством воды.К ним относятся:
- Повышенная неопределенность в местах и масштабах, которые отличаются от существующих данных мониторинга, используемых для калибровки модели
- Возможность получения разных результатов моделирования для моделей, разработанных в разных масштабах
В первом случае прогнозы SPARROW потенциально могут иметь более высокую неопределенность для размеров водосборных бассейнов, меньших или больших, чем у участков мониторинга водотоков, используемых для калибровки модели. Одной из проблем для модели SPARROW, как и для любой модели водосбора, является сложность получения большой выборки участков мониторинга из небольших водосборных бассейнов, где условия окружающей среды и соответствующее качество речной воды могут значительно различаться. Из-за неоднородности условий в малых масштабах и ограниченных ресурсов мониторинга в этих пространственных масштабах доступно меньше данных мониторинга водотоков. Для национальных моделей SPARROW размеры контролируемых водосборных бассейнов варьируются от нескольких сотен квадратных километров до значительно более 1000 квадратных километров, тогда как более современные региональные модели SPARROW обычно имеют гораздо больше участков мониторинга, расположенных в меньших водоразделах (более новые национальные версии модели SPARROW). которые находятся в стадии разработки, будут использовать эти более подробные данные мониторинга).Например, в модели общего азота SAGT (Южная Атлантика-Залив Теннесси) SPARROW (Hoos and McMahon, 2009) контролируемые водосборные бассейны имеют размер от 15 до 28 000 квадратных километров (межквартильный диапазон от 230 до 1600 квадратных километров). Тем не менее, важно признать, что как в национальной, так и в региональной моделях площади дополнительных дренажей (т.
Из-за неоднородности условий в малых масштабах и ограниченных ресурсов мониторинга в этих пространственных масштабах доступно меньше данных мониторинга водотоков. Для национальных моделей SPARROW размеры контролируемых водосборных бассейнов варьируются от нескольких сотен квадратных километров до значительно более 1000 квадратных километров, тогда как более современные региональные модели SPARROW обычно имеют гораздо больше участков мониторинга, расположенных в меньших водоразделах (более новые национальные версии модели SPARROW). которые находятся в стадии разработки, будут использовать эти более подробные данные мониторинга).Например, в модели общего азота SAGT (Южная Атлантика-Залив Теннесси) SPARROW (Hoos and McMahon, 2009) контролируемые водосборные бассейны имеют размер от 15 до 28 000 квадратных километров (межквартильный диапазон от 230 до 1600 квадратных километров). Тем не менее, важно признать, что как в национальной, так и в региональной моделях площади дополнительных дренажей (т. е. площади, расположенные между вложенными участками мониторинга), которые обычно меньше, чем площади, отраженные в распределении размеров общего дренажа для участков мониторинга. , предоставить дополнительную мелкомасштабную информацию, которая поможет в оценке процессов снабжения и переноса питательных веществ при калибровке модели.
е. площади, расположенные между вложенными участками мониторинга), которые обычно меньше, чем площади, отраженные в распределении размеров общего дренажа для участков мониторинга. , предоставить дополнительную мелкомасштабную информацию, которая поможет в оценке процессов снабжения и переноса питательных веществ при калибровке модели.
Повышенная неопределенность может также возникать для прогнозов модели в меньших пространственных масштабах, поскольку некоторые наборы данных, используемые для калибровки моделей SPARROW, сообщаются для более крупных площадей, чем площади водосбора, для которых рассчитываются прогнозы модели или где осуществляется мониторинг. Например, отчетность на уровне уезда (межквартильный диапазон = от 1000 до 2500 квадратных километров) является обычной для многих государственных сельскохозяйственных баз данных (например, по внесению удобрений, отходам животноводства), хотя пространственная точность данных на уровне уезда часто повышается. с помощью данных о землепользовании (т. g., разрешение 30 м или 1 км) в моделировании SPARROW для пространственного уточнения местоположения сельскохозяйственной деятельности в округах.
g., разрешение 30 м или 1 км) в моделировании SPARROW для пространственного уточнения местоположения сельскохозяйственной деятельности в округах.
SPARROW предоставляет инструмент для интеграции доступной информации о водных ресурсах в различных пространственных масштабах. Доверительные интервалы, представленные для прогнозов модели SPARROW (т. е. нагрузки, выходы, доли источников), включают наблюдаемую изменчивость в работе модели в диапазоне данных калибровки и, таким образом, обеспечивают надежную меру статистической неопределенности в сообщаемых прогнозах. .В пространственных масштабах (как правило, небольших водосборах), которые отличаются от масштабов, отражаемых геопространственными данными, используемыми для калибровки модели, неопределенность прогнозов модели потенциально больше, чем сообщаемые неопределенности. Из-за широкого диапазона изменчивости условий качества речной воды в небольших пространственных масштабах не существует установленных рекомендаций или правил относительно величины неопределенностей в прогнозах SPARROW, когда модель применяется за пределами диапазона калибровки. Тем не менее, данные мониторинга и геопространственные данные, используемые для калибровки, могут дать приблизительное представление о пространственных пределах предсказаний модели.Таким образом, рекомендуется, чтобы пользователи модели признавали, что сообщаемые меры неопределенности дают лишь приблизительные оценки надежности прогнозов в случаях, когда модель применяется за пределами диапазона калибровки.
Тем не менее, данные мониторинга и геопространственные данные, используемые для калибровки, могут дать приблизительное представление о пространственных пределах предсказаний модели.Таким образом, рекомендуется, чтобы пользователи модели признавали, что сообщаемые меры неопределенности дают лишь приблизительные оценки надежности прогнозов в случаях, когда модель применяется за пределами диапазона калибровки.
Второе ограничение прогнозов SPARROW, связанное с масштабом, заключается в том, что модели, откалиброванные для разных, но перекрывающихся пространственных областей, могут не давать идентичных прогнозов средних условий (например, нагрузки, выработки, долей источников) для данного водосбора, даже если этот водосбор является подмножеством большей площади водосбора, включенной в одну из моделей.Это можно объяснить различиями в калибровочных данных, которые могут отображать разные диапазоны изменчивости условий окружающей среды в разных пространственных масштабах. Такие различия могут привести к тому, что оценочные параметры модели и прогнозы будут различаться между моделями SPARROW. Воздействие экологических процессов и свойств на условия качества речной воды также может различаться по своей важности в различных пространственных масштабах, особенно в отношении многолетних среднегодовых условий, отраженных в моделях SPARROW.Например, долгосрочная среднегодовая температура может иметь важное значение для объяснения изменений среднегодовой доставки питательных веществ в Соединенных Штатах или в пределах большого региона Соединенных Штатов; это может соответствовать различиям в реакции качества воды на большие различия температур в одном штате (например, в Северной Каролине) по сравнению с другими (Вайоминг). Тем не менее, температура может мало объяснить различия в доставке питательных веществ в пределах одного штата из-за небольшой пространственной изменчивости среднегодовых температур.В таких случаях региональная модель SPARROW может дать более широкое и более точное представление о факторах, влияющих на пространственную изменчивость долгосрочных среднегодовых условий качества воды в реке, чем может дать мелкомасштабная пространственная модель.
Воздействие экологических процессов и свойств на условия качества речной воды также может различаться по своей важности в различных пространственных масштабах, особенно в отношении многолетних среднегодовых условий, отраженных в моделях SPARROW.Например, долгосрочная среднегодовая температура может иметь важное значение для объяснения изменений среднегодовой доставки питательных веществ в Соединенных Штатах или в пределах большого региона Соединенных Штатов; это может соответствовать различиям в реакции качества воды на большие различия температур в одном штате (например, в Северной Каролине) по сравнению с другими (Вайоминг). Тем не менее, температура может мало объяснить различия в доставке питательных веществ в пределах одного штата из-за небольшой пространственной изменчивости среднегодовых температур.В таких случаях региональная модель SPARROW может дать более широкое и более точное представление о факторах, влияющих на пространственную изменчивость долгосрочных среднегодовых условий качества воды в реке, чем может дать мелкомасштабная пространственная модель. Как правило, результаты региональной модели могут применяться в конкретном субрегионе (например, в штате) с теми же ограничениями неопределенности, которые действуют на всей протяженности модели (например, что отражено в доверительных интервалах прогнозов), если только не определено местные условия окружающей среды и деятельность человека, которые, как известно, влияют на качество воды в этом субрегионе, не представлены в региональной модели, или если пространственная структура в структуре остатков (т.е., ошибки прогноза) в этом субрегионе позволяют предположить, что некоторые факторы, влияющие на качество воды, не представлены в региональной модели. Таким образом, пользователям результатов моделирования важно осознавать эти ограничения и выбирать результаты из моделей SPARROW, которые представляют пространственные масштабы для их приложений управления.
Как правило, результаты региональной модели могут применяться в конкретном субрегионе (например, в штате) с теми же ограничениями неопределенности, которые действуют на всей протяженности модели (например, что отражено в доверительных интервалах прогнозов), если только не определено местные условия окружающей среды и деятельность человека, которые, как известно, влияют на качество воды в этом субрегионе, не представлены в региональной модели, или если пространственная структура в структуре остатков (т.е., ошибки прогноза) в этом субрегионе позволяют предположить, что некоторые факторы, влияющие на качество воды, не представлены в региональной модели. Таким образом, пользователям результатов моделирования важно осознавать эти ограничения и выбирать результаты из моделей SPARROW, которые представляют пространственные масштабы для их приложений управления.
Что вызывает неопределенность в моделях SPRROW?
Все модели, включая ВОРОБЬЯ, являются несовершенными представлениями реальности и, следовательно, связаны с неопределенностью. Есть много причин для этой неопределенности, в том числе:
Есть много причин для этой неопределенности, в том числе:
- Ограничения в поддержке мониторинга потоков и геопространственных данных
- Ограничения в понимании экологических процессов, влияющих на качество воды
- Ограничения подхода моделирования в точном представлении процессов окружающей среды
Трудно точно определить количество неопределенности, связанной с двумя последними пунктами. Однако неопределенность, вызванная ограничениями в базовых наборах данных, понимается и постоянно улучшается.
Одной из важных причин неопределенности является ограничение количества мест мониторинга, доступных для калибровки модели. Как и в любой статистической модели, неопределенность в моделях SPARROW уменьшается по мере увеличения числа участков, доступных для калибровки. В случае СПАРРОУ количество калибровочных участков определяется количеством участков с достаточным качеством воды и данными о расходах для расчета составляющей нагрузки. Исторически количество сайтов, представленных в программах мониторинга федеральных, государственных и местных агентств, колебалось. Однако в последнее время количество участков с достаточными данными мониторинга стало сокращаться. Следует ожидать, что такое снижение вызовет большую неопределенность во всех экологических моделях, включая SPRROW.
Однако в последнее время количество участков с достаточными данными мониторинга стало сокращаться. Следует ожидать, что такое снижение вызовет большую неопределенность во всех экологических моделях, включая SPRROW.
Неопределенность в моделях SPARROW также определяется качеством геопространственных данных, доступных для построения данных независимых переменных. Многие наборы данных, из которых получаются важные переменные, доступны только в относительно грубом масштабе, например, для округов. Для разработки прогнозных данных из таких наборов данных значения должны быть распределены по меньшему масштабу водосборных бассейнов.Это приводит к тому, что данные предиктора имеют пространственную неопределенность, хотя эта неопределенность обычно отражается в ошибках модели в процессе калибровки.
Как учитывается неопределенность в моделях SPARROW?
Неопределенность всегда присутствует в моделях окружающей среды, таких как ВОРБЕЙ. Неопределенность может быть вызвана многими факторами, но часто она связана с ограничениями количества и качества наборов вспомогательных данных. Эти ограничения неизбежны из-за масштабности усилий и нехватки ресурсов для поддержки более обширной разработки базы данных.Например, данные о качестве воды не могут постоянно собираться в контролируемых местах ручья. Таким образом, существуют внутренние неопределенности измерений при описании нагрузок на качество речной воды по данным мониторинга, которые используются для калибровки модели SPARROW.
Эти ограничения неизбежны из-за масштабности усилий и нехватки ресурсов для поддержки более обширной разработки базы данных.Например, данные о качестве воды не могут постоянно собираться в контролируемых местах ручья. Таким образом, существуют внутренние неопределенности измерений при описании нагрузок на качество речной воды по данным мониторинга, которые используются для калибровки модели SPARROW.
Как и все статистические модели, модели SPARROW разрабатываются посредством процесса калибровки, в ходе которого оцениваются значения параметров, чтобы свести к минимуму неопределенность при прогнозировании нагрузок на составляющие потоки. Это достигается с помощью статистического алгоритма, называемого нелинейным методом наименьших квадратов, в котором значения параметров оцениваются для минимизации квадрата разницы между измеренной и прогнозируемой нагрузками.Неопределенность количественно определяется как остаточная ошибка в прогнозировании нагрузки, которая не может быть учтена посредством настройки параметров. Общая неопределенность в модели количественно определяется с помощью ряда статистических диагностических методов. Однако наиболее распространенная мера неопределенности называется «среднеквадратической ошибкой», которая представляет собой просто среднее квадратов разностей между измеренной и прогнозируемой нагрузками.
Общая неопределенность в модели количественно определяется с помощью ряда статистических диагностических методов. Однако наиболее распространенная мера неопределенности называется «среднеквадратической ошибкой», которая представляет собой просто среднее квадратов разностей между измеренной и прогнозируемой нагрузками.
Как можно использовать неопределенность модели SPARROW для лучшего понимания факторов, влияющих на качество воды?
Модели SPARROW предназначены для учета пространственной изменчивости данных мониторинга качества речной воды путем связывания их с пространственно определенными факторами окружающей среды.Неопределенность в моделях количественно определяется различиями между измеренными и смоделированными оценками нагрузки на реку, обычно называемыми остатками. Процесс калибровки предназначен для минимизации этих различий. Меры неопределенности используются для оценки качества соответствия модели и используются для оценки предела погрешности в прогнозировании нагрузок, концентраций, выходов и вкладов источников.
Пространственные модели высоких или низких невязок, которые можно наблюдать на картах, потенциально могут выявить важные факторы окружающей среды, которые могут не учитываться в модели.Например, модель SPARROW может постоянно завышать нагрузки азотом в определенной части модельного региона. Эти большие невязки могут выявить пространственный паттерн, совпадающий с характеристикой водосбора, которая может быть связана с ними. В случае азота, например, большие остатки могут возникать там, где важна денитрификация. Денитрификация может быть усилена в районах с особыми почвенными или геологическими характеристиками, такими как высокая проницаемость почвы. Характер остатков модели может указывать на то, что включение таких объясняющих переменных важно.Таким образом, визуализация карт пространственных закономерностей в неопределенности модели SPARROW обычно используется для исследования (и учета в последующих калибровках модели) факторов окружающей среды, которые могут быть связаны с важными глубинными процессами, влияющими на качество воды.
Моделирование СПАРРОУ: последствия для управления и мониторинга
Как можно использовать модели SPARROW для планирования будущих программ мониторинга?
Модели SPARROW носят статистический характер, и их неопределенность часто зависит как от качества, так и от количества данных, доступных для калибровки.Для моделей SPARROW данные калибровки состоят из оценок нагрузки на участках мониторинга. Неточные или неточные измерения нагрузки на объектах мониторинга создадут неопределенность в моделях, равно как и меньшее количество объектов мониторинга. Там, где неопределенность связана с большими ошибками прогнозирования, потенциально может быть реализован дополнительный или уточненный мониторинг для уменьшения неопределенности. В более общем смысле, сбор данных для калибровки SPARROW может выявить ограничения в доступных данных мониторинга нагрузки. Эта информация может помочь агентствам сделать их программы мониторинга более эффективными.
SPARROW нетипичны для моделирования качества воды, поскольку они могут помочь в интерпретации данных, собранных в сети пунктов мониторинга (Smith and others, 1997). После создания модели SPARROW для сети мониторинга существует некоторая общность между целями мониторинга и целями моделирования SPARROW. Тогда становится логичным рассмотреть возможность использования модели для выбора мест отбора проб, которые одновременно оптимизируют цели мониторинга и моделирования.
Чтобы спроектировать оптимальную сеть мониторинга, необходимо сначала четко определить цель(и) мониторинга. Затем модель SPARROW может служить инфраструктурой алгоритма оптимизации проектирования сети. Например, МакМахон и другие (2003 г.) предложили использовать модель SPARROW для обнаружения нового мониторинга, чтобы обеспечить наиболее точные основанные на модели прогнозы превышения критерия качества воды в реке. Для этой цели имеет смысл собирать данные в местах водотоков, где предсказание превышения моделью очень близко к пороговому значению и в то же время весьма неопределенно (вместо того, чтобы сосредоточиться на местах водотоков, где предсказание превышает пороговое значение очень часто или редко). Модели SPARROW могут предоставить количественную информацию о неопределенности предсказания модели для каждого участка водотока и, таким образом, указать, где дополнительная выборка лучше всего поддержит цель. Цели могут также касаться других аспектов неопределенности модели.
Модели SPARROW могут предоставить количественную информацию о неопределенности предсказания модели для каждого участка водотока и, таким образом, указать, где дополнительная выборка лучше всего поддержит цель. Цели могут также касаться других аспектов неопределенности модели.
Как выводы могут быть использованы лицами, принимающими решения, и заинтересованными сторонами?
Моделирование SPARROW и интерпретация результатов моделирования расширяют наше понимание пространственных различий в источниках загрязнения и экологических и гидрологических процессах, которые контролируют их перенос и доставку в воды нижнего течения.Выводы имеют важное значение для разработки политики и управления, связанных с проблемами качества воды. Прогнозы моделей и связанная с ними информация могут помочь штатам, федеральным агентствам и другим заинтересованным сторонам определить вклад питательных веществ в водотоки из источников загрязнения и стоков вверх по течению (например, суббассейны, штаты).
Одним из применений результатов SPARROW, которые могут помочь лучшему пониманию биогенных нагрузок и источников водотока, является их использование в оценках общей максимальной суточной нагрузки (TMDL), особенно в водосборных бассейнах, где собраны небольшие или ограниченные данные мониторинга водотока.Например, результаты SPARROW для региона Новой Англии ранее использовались в оценке TMDL нагрузки азотом от реки Коннектикут до пролива Лонг-Айленд для решения проблем, связанных с сезонной прибрежной гипоксией и расположением источников водосборных бассейнов. В этом примере SPRROW предоставил информацию (Moore and others, 2004) о поступлении азота в пролив из точечных и неточечных источников азота (например, атмосферные отложения, муниципальные сточные воды) из водосборных бассейнов вверх по течению в четырех штатах и суббассейнах Коннектикута. Бассейн реки.
SPARROW также предоставляет информацию, которую можно использовать для нацеливания на водосборные бассейны, расположенные выше по течению, где можно ожидать, что реализация стратегий управления приведет к более эффективному сокращению биогенных нагрузок в водоемах ниже по течению на единицу усилий, затрачиваемых вверх по течению. Эти водосборные бассейны включают в себя те, где прогнозы SPRROW по «доставленному урожаю» — массе питательных веществ на единицу площади, доставляемой в воды нижнего течения, — высоки. Водосборные бассейны с высокой урожайностью обычно имеют большое количество питательных веществ из точечных и / или неточечных источников; эти водосборные бассейны также эффективно доставляют питательные вещества в воды нижнего течения, потому что, как правило, меньшее количество питательных веществ удаляется при транспортировке по суше и в воде по сравнению с водосборными бассейнами с низким уровнем доставки.Этот тип информации SPARROW использовался штатами и федеральными агентствами для определения водоразделов вверх по течению, которые должны получить приоритетное управление. Например, Департамент природных ресурсов Мэриленда использует оценки обеспеченного урожая (и связанных с ним долей источников для сельского хозяйства, атмосферных отложений и т. д.) с помощью SPARROW, чтобы помочь в целенаправленных усилиях по сокращению биогенных веществ в водосборных бассейнах Чесапикского залива.
Эти водосборные бассейны включают в себя те, где прогнозы SPRROW по «доставленному урожаю» — массе питательных веществ на единицу площади, доставляемой в воды нижнего течения, — высоки. Водосборные бассейны с высокой урожайностью обычно имеют большое количество питательных веществ из точечных и / или неточечных источников; эти водосборные бассейны также эффективно доставляют питательные вещества в воды нижнего течения, потому что, как правило, меньшее количество питательных веществ удаляется при транспортировке по суше и в воде по сравнению с водосборными бассейнами с низким уровнем доставки.Этот тип информации SPARROW использовался штатами и федеральными агентствами для определения водоразделов вверх по течению, которые должны получить приоритетное управление. Например, Департамент природных ресурсов Мэриленда использует оценки обеспеченного урожая (и связанных с ним долей источников для сельского хозяйства, атмосферных отложений и т. д.) с помощью SPARROW, чтобы помочь в целенаправленных усилиях по сокращению биогенных веществ в водосборных бассейнах Чесапикского залива.
Министерство здравоохранения и окружающей среды Канзаса также использовало прогнозы SPARROW о доставке питательных веществ в Мексиканский залив для определения водосборных бассейнов внутри штата, которым следует уделять первоочередное внимание при управлении питательными веществами.Это часть усилий штата Канзас по смягчению последствий чрезмерного обогащения питательными веществами и эвтрофикации в водах штата и прибрежных водах Мексиканского залива (см. «Канзасский план сокращения содержания питательных веществ», 2004 г.).
Кроме того, Министерство сельского хозяйства США недавно рекомендовало использовать результаты SPARROW вместе с другой информацией об источниках питательных веществ, чтобы нацелить размещение методов сохранения в 12 штатах в бассейне реки Миссисипи, чтобы контролировать поступление питательных веществ вниз по течению в Мексиканский залив. (т.е.g., см. Инициатива USDA Healthy Watershed Initiative (2009 г.)).
Прогнозы речных нагрузок, источников питательных веществ и переноса с помощью SPARROW также могут предоставить широкомасштабную информацию в поддержку региональных оценок источников питательных веществ в прибрежных и внутренних водах. Например, ранее NOAA использовало результаты SPARROW для помощи в оценке подверженности эстуариев в континентальных Соединенных Штатах обогащению питательными веществами (Национальное исследование эвтрофикации NOAA, 1999). Служба охраны природных ресурсов Министерства сельского хозяйства США (NRCS) также использовала SPARROW для информирования о калибровке и проверке их применения моделей сельскохозяйственных водосборных бассейнов в рамках оценки воздействия методов сохранения на качество воды в стране в рамках Программы оценки и оценки сохранения (CEAP). .
Например, ранее NOAA использовало результаты SPARROW для помощи в оценке подверженности эстуариев в континентальных Соединенных Штатах обогащению питательными веществами (Национальное исследование эвтрофикации NOAA, 1999). Служба охраны природных ресурсов Министерства сельского хозяйства США (NRCS) также использовала SPARROW для информирования о калибровке и проверке их применения моделей сельскохозяйственных водосборных бассейнов в рамках оценки воздействия методов сохранения на качество воды в стране в рамках Программы оценки и оценки сохранения (CEAP). .
SPARROW также могут информировать политиков по вопросам управления питательными веществами и установления критериев питательных веществ. В 2009 году группа Национального исследовательского совета назвала SPARROW важным информационным инструментом, который может помочь в определении приоритетных водосборных бассейнов для целенаправленного управления и контроля питательных веществ в бассейне реки Миссисипи. Выводы SPARROW также улучшили понимание влияния верховьев воды на качество воды ниже по течению (например, Александр и другие, 2007 г. ), что способствовало лучшему пониманию юрисдикции федерального Закона о чистой воде и рекомендаций для регионов АООС США и США.S. Армейский корпус инженерных округов (например, см. ссылку на сборник Журнала Американской ассоциации водных ресурсов, посвященный гидрологии верховьев.
), что способствовало лучшему пониманию юрисдикции федерального Закона о чистой воде и рекомендаций для регионов АООС США и США.S. Армейский корпус инженерных округов (например, см. ссылку на сборник Журнала Американской ассоциации водных ресурсов, посвященный гидрологии верховьев.
Каково значение результатов SPARROW для моделирования и мониторинга?
Разработка гидрологических моделей и моделей качества воды значительно продвинулась за последние 20 лет, что привело к улучшению всесторонних оценок условий качества воды и улучшению понимания ключевых факторов и процессов, влияющих на качество воды, таких как землепользование, химические источники загрязнения, особенности природного ландшафта и гидрологический перенос.
- Успех подхода модели SPARROW зависит от: Точной и подробной пространственной информации о водосборном бассейне, включая информацию о структуре посевов, городском населении, сбросах из точечных источников и использовании навоза
- Пространственно обширные долгосрочные данные о качестве воды в сочетании с данными о речном стоке
- Непрерывные исследования и применение моделей, которые явно учитывают процессы на суше и воде и то, как они определяют перемещение загрязняющих веществ вниз по течению
К сожалению, мониторинг качества воды со стороны федеральных агентств и агентств штата заметно сократился. Например, в 1975-1980 годах, когда мониторинг был на пике, количество мест в США, где Геологическая служба США собирала данные о питательных веществах, подходящие для использования в таких исследованиях, как модель SPARROW, или для анализа долгосрочных тенденций, составляло около 4500. Напротив, количество станций, которое мы имеем сегодня, составляет всего около 1900 станций, что составляет снижение примерно на 60 процентов.
Например, в 1975-1980 годах, когда мониторинг был на пике, количество мест в США, где Геологическая служба США собирала данные о питательных веществах, подходящие для использования в таких исследованиях, как модель SPARROW, или для анализа долгосрочных тенденций, составляло около 4500. Напротив, количество станций, которое мы имеем сегодня, составляет всего около 1900 станций, что составляет снижение примерно на 60 процентов.
В дополнение к мониторингу качества воды отсутствует большая часть дополнительных пространственных данных, необходимых для интерпретации данных о качестве воды, включая более точную информацию о сбросах из точечных источников, использовании химикатов, изменениях в землепользовании, водопользовании, методах управления земельными ресурсами. , усилия по сохранению, геоморфология и сети потоков, а также геологические условия.В настоящее время мы сталкиваемся с ограничениями данных в отношении вспомогательной информации, необходимой для поддержки моделей, включая информацию о точечных источниках, сельскохозяйственных применениях и методах, а также об изменениях в методах управления земельными ресурсами (в том числе, например, о лучших усилиях по управлению и сохранению).
Мы должны продолжать интегрировать долгосрочный наземный мониторинг с прогностическими инструментами, чтобы обеспечить соответствующее представление физических, химических и биологических процессов в моделях, в сочетании с мощными статистическими методами для оценки важности различных факторов, используемых в модели.Непрерывный мониторинг и сбор данных снизят общую неопределенность прогнозов и оценок моделей. В свою очередь, анализ неопределенности, связанный с каждым прогнозом, поможет определить будущие потребности в мониторинге и сборе данных.
ВОРОБЕЙ Моделирование в Южной Атлантике-регион Теннесси
В каком масштабе результаты модели SAGT SPARROW применимы во Флориде?
Было высказано предположение, что модель общего содержания азота в Южной Атлантике и заливе Теннесси (SAGT)-SPARROW является региональной моделью и не откалибрована для Флориды.Модели SPARROW, как правило, разрабатываются так, чтобы они не зависели от масштаба, и считаются действительными в широком диапазоне пространственных масштабов, что приблизительно соответствует масштабу геопространственных данных, используемых для калибровки модели. Как правило, модели SPARROW можно использовать для моделирования условий качества воды в масштабах, варьирующихся от водосборных бассейнов одного ручья (от нескольких квадратных километров до десятков квадратных километров) до размеров крупных водоразделов и речных бассейнов, охватывающих тысячи квадратных километров. .Таким образом, модели SPARROW являются гибкими и могут использоваться для выполнения различных типов оценок, охватывающих диапазон размеров бассейнов.
Как правило, модели SPARROW можно использовать для моделирования условий качества воды в масштабах, варьирующихся от водосборных бассейнов одного ручья (от нескольких квадратных километров до десятков квадратных километров) до размеров крупных водоразделов и речных бассейнов, охватывающих тысячи квадратных километров. .Таким образом, модели SPARROW являются гибкими и могут использоваться для выполнения различных типов оценок, охватывающих диапазон размеров бассейнов.
Геопространственные данные регионального масштаба, полученные вдоль границ водоразделов, использовались для калибровки модели общего азота SAGT-SPARROW (Hoos and McMahon, 2009). Способность модели точно прогнозировать нагрузки в малых пространственных масштабах ограничена степенью пространственного разрешения в наборах геопространственных данных. Некоторые из этих наборов геопространственных данных, например данные об использовании удобрений и животноводстве, разрешаются только на уровне округа, хотя пространственная точность данных на уровне округа была повышена для модели общего азота SAGT SPARROW за счет использования подробных данных о землепользовании. (Набор данных о национальном земельном покрове Геологической службы США) для пространственного уточнения местоположения сельскохозяйственной деятельности в округах.Тем не менее, поскольку средний размер округа в области модели SAGT SPARROW составляет 1425 км 90 317 2 90 318 , мы предложили проявлять особую осторожность при интерпретации условий источника азота для участков, где размер водосбора примерно такой же или меньше. Таким образом, прогнозы SAGT-SPARROW по общей азотной нагрузке и долям источников могут использоваться для оценки условий качества воды на конечных участках водосборных бассейнов, вносящих вклад, например, в залив Чоктаватчи или гавань Шарлотт (площадь водосборного бассейна = 13 496 и 8 134 км 2 , соответственно), но его следует использовать с осторожностью для оценки условий на конечных участках водоразделов, участвующих, например, в заливе Сарасота (653 км 2 ).
(Набор данных о национальном земельном покрове Геологической службы США) для пространственного уточнения местоположения сельскохозяйственной деятельности в округах.Тем не менее, поскольку средний размер округа в области модели SAGT SPARROW составляет 1425 км 90 317 2 90 318 , мы предложили проявлять особую осторожность при интерпретации условий источника азота для участков, где размер водосбора примерно такой же или меньше. Таким образом, прогнозы SAGT-SPARROW по общей азотной нагрузке и долям источников могут использоваться для оценки условий качества воды на конечных участках водосборных бассейнов, вносящих вклад, например, в залив Чоктаватчи или гавань Шарлотт (площадь водосборного бассейна = 13 496 и 8 134 км 2 , соответственно), но его следует использовать с осторожностью для оценки условий на конечных участках водоразделов, участвующих, например, в заливе Сарасота (653 км 2 ).
Модель общего азота SAGT-SPARROW была откалибрована, чтобы соответствовать наблюдаемым измерениям потоков питательных веществ на 321 участке мониторинга площадью от 15 до 28 000 квадратных километров (межквартильный диапазон от 230 до 1600 квадратных километров). В остатках модели не наблюдалось пространственной структуры, что указывает на то, что модель работала лучше или хуже во Флориде по сравнению с другими штатами или водоразделами в области модели SAGT.
В остатках модели не наблюдалось пространственной структуры, что указывает на то, что модель работала лучше или хуже во Флориде по сравнению с другими штатами или водоразделами в области модели SAGT.
Геология в некоторых частях Флориды способствует обмену воды и азота между поверхностными и грунтовыми водами.Действует ли SPARROW в этих областях?
Как обсуждалось в Hoos and McMahon (2009), подход SPARROW к объяснению нагрузок в русле на основе атрибутов водосбора имеет ограничения для участков водотока, для которых фактические гидрологические границы не соответствуют кажущемуся водоразделу, определенному по топографии поверхности. Это может происходить в некоторых речных бассейнах в северной и центральной Флориде, где обмен стока с нижележащим региональным водоносным горизонтом способствует значительному притоку азота в бассейны поверхностных вод и оттоку из них.Модельные прогнозы представлены для речных бассейнов, отнесенных к этой проблеме (бассейны рек Оклаваха, Кристал, Нижний Санте-Фе, Нижний Сувани, Сент-Маркс и Чипола), но они могут быть менее надежными из-за этого немоделируемого компонента потока. Кроме того, на точность прогнозов модели SPARROW для рек Сент-Джонс и Индиан-Ривер может повлиять неадекватное представление гидрологической сети цифровой сегментированной сетью ERF1_2, используемой в модели.
Кроме того, на точность прогнозов модели SPARROW для рек Сент-Джонс и Индиан-Ривер может повлиять неадекватное представление гидрологической сети цифровой сегментированной сетью ERF1_2, используемой в модели.
Дэвид Воробей | Медицинский факультет
Келли Р.С., Стюарт И.Д., Бейн Х., Кахру П., Спиро Иии А., Воконас П., Воробей Д., Вайс С.Т., Книхтиля Х.М., Литоньюа А.А., Уэрхэм Н.Дж., Лангенберг С., Ласки-Су Дж.А.Метаболомические различия в показателях функции легких: данные двух когорт. грудная клетка. 14 октября 2021 г. PMID: 34650005.
Читать в: PubMedКелли Р.С., Бейн Х., Спиро А., Воконас П., Воробей Д., Вайс С.Т., Шварц Дж., Нассан Ф.Л., Ли-Сарвар К., Хуанг М., Качроо П., Чу С.Х., Литонжуа А.А., Ласки-Су Дж.А. Метаболомические признаки воздействия свинца в исследовании нормативного старения VA. Окружающая среда Рез. 2020 11; 190:110022. PMID: 327
.
Читать в: PubMed 08.2020
08.2020Брин М., Нванаджи-Энверем Дж. К., Карраш С., Флекседер С., Шульц Х., Вальденбергер М., Кунце С., Оллерт М., Вайдингер С., Колицино Э. , Гао Х, Ван С, Шен Дж, Джаст А.С., Воконас П, Воробей Д, Хоу Л, Шварц ДД, Баккарелли АА, Петерс А, Уорд-Кавинесс КК.Ускоренное эпигенетическое старение как фактор риска хронической обструктивной болезни легких и снижение функции легких в двух проспективных когортных исследованиях. Старение (Олбани, штат Нью-Йорк). 2020 08 03; 12(16):16539-16554. PMID: 32747609.
Читать в: PubMedGao X, Coull B, Lin X, Vokonas P, Sparrow D, Hou L, DeMeo DL, Litonjua AA, Schwartz J, Baccarelli AA. Связь соотношения нейтрофилов и лимфоцитов с функцией легких в 30-летнем лонгитюдном исследовании ветеранов США.JAMA Сеть открыта. 2020 07 01; 3(7):e2010350. PMID: 32658288.
Читать в: PubMed Wang C, Just A, Heiss J, Coull BA, Hou L, Zheng Y, Sparrow D, Vokonas PS, Baccarelli A, Schwartz J. Биомаркеры старения и функции легких в нормативном исследовании старения. Старение (Олбани, штат Нью-Йорк). 2020 06 19; 12(12):11942-11966. PMID: 32561690.
Биомаркеры старения и функции легких в нормативном исследовании старения. Старение (Олбани, штат Нью-Йорк). 2020 06 19; 12(12):11942-11966. PMID: 32561690.
Wang X, Kim D, Tucker KL, Weisskopf MG, Sparrow D, Hu H, Park SK. Влияние потребления натрия и калия с пищей на мобилизацию костного свинца среди мужчин среднего и пожилого возраста: исследование нормативного старения по делам ветеранов.Питательные вещества. 2019 13 ноября; 11(11). PMID: 31766133.
Читать в: PubMedColicino E, Just A, Kioumourtzoglou MA, Vokonas P, Cardenas A, Sparrow D, Weisskopf M, Nie LH, Hu H, Schwartz JD, Wright Р.О., Баккарелли А.А. Биомаркеры метилирования ДНК крови кумулятивного воздействия свинца у взрослых. J Expo Sci Environ Epidemiol. 2021 02; 31(1):108-116. PMID: 31636367.
Читать в: PubMed Shrine N, Guyatt AL, Erzurumluoglu AM, Jackson VE, Hobbs BD, Melbourne CA, Batini C, Fawcett KA, Song K, Sakornsakolpat P, Li X, Боксалл Р. , Рив Н.Ф., Обейдат М., Чжао Дж. Х., Вильшер М., Вайс С., Кентисту К. А., Кук Дж. П., Сун Б. Б., Чжоу Дж., Хуэй Дж., Карраш С., Имбоден М., Харрис С. Э., Мартен Дж., Энрот С., Керр С.М., Суракка И., Витарт В., Лехтимяки Т., Аллен Р.Дж., Бакке П.С., Бити Т.Х., Бликер Э.Р., Боссе И., Брандсма К.А., Чен З., Крапо Д.Д., Данеш Дж., ДеМео Д.Л., Дадбридж Ф., Эверт Р., Гигер С. , Гулсвик А., Ханселл А.Л., Хао К., Хоффман Дж.Д., Хокансон Дж.Е., Хомут Г., Джоши П.К., Жубер П., Лангенберг С., Ли Х, Ли Л., Лин К., Линд Л., Локанторе Н., Луан Дж., Махаджан А., Маранвиль JC, Мюррей А., Никл Д.С., Пакер Р., Паркер М.М., Пейнтон М.Л., Портеус Д.Дж., Прокопенко Д., Цяо Д., Равал Р., Рунц Х., Сайерс И., Син Д.Д., Смит Б.Х., Артигас М.С., Воробей Д., Тал-Сингер R, Timmers PRHJ, Van den Berge M, Whittaker JC, Woodruff PG, Yerges-Armstrong LM, Troyanskaya OG, Raitakari OT , Кяхёнен М., Полашек О., Гилленстен У., Рудан И., Дири И.Дж., Пробст-Хенш Н.М., Шульц Х., Джеймс А.Л., Уилсон Дж.Ф., Стуббе Б., Зеггини Э.
, Рив Н.Ф., Обейдат М., Чжао Дж. Х., Вильшер М., Вайс С., Кентисту К. А., Кук Дж. П., Сун Б. Б., Чжоу Дж., Хуэй Дж., Карраш С., Имбоден М., Харрис С. Э., Мартен Дж., Энрот С., Керр С.М., Суракка И., Витарт В., Лехтимяки Т., Аллен Р.Дж., Бакке П.С., Бити Т.Х., Бликер Э.Р., Боссе И., Брандсма К.А., Чен З., Крапо Д.Д., Данеш Дж., ДеМео Д.Л., Дадбридж Ф., Эверт Р., Гигер С. , Гулсвик А., Ханселл А.Л., Хао К., Хоффман Дж.Д., Хокансон Дж.Е., Хомут Г., Джоши П.К., Жубер П., Лангенберг С., Ли Х, Ли Л., Лин К., Линд Л., Локанторе Н., Луан Дж., Махаджан А., Маранвиль JC, Мюррей А., Никл Д.С., Пакер Р., Паркер М.М., Пейнтон М.Л., Портеус Д.Дж., Прокопенко Д., Цяо Д., Равал Р., Рунц Х., Сайерс И., Син Д.Д., Смит Б.Х., Артигас М.С., Воробей Д., Тал-Сингер R, Timmers PRHJ, Van den Berge M, Whittaker JC, Woodruff PG, Yerges-Armstrong LM, Troyanskaya OG, Raitakari OT , Кяхёнен М., Полашек О., Гилленстен У., Рудан И., Дири И.Дж., Пробст-Хенш Н.М., Шульц Х., Джеймс А.Л., Уилсон Дж.Ф., Стуббе Б., Зеггини Э. , Ярвелин М.Р., Уэрхэм Н., Сильверман Э.К., Хейуорд С., Моррис А.П. , Баттерворт А.С., Скотт Р.А., Уолтерс Р.Г., Мейерс Д.А., Чо М.Х., Страчан Д.П., Холл И.П., Тобин М.Д., Уэйн Л.В.Исправление автора: новые генетические сигналы для функции легких выявляют пути и ассоциации хронической обструктивной болезни легких у разных предков. Нат Жене. 2019 июнь; 51(6):1067. PMID: 31110354.
, Ярвелин М.Р., Уэрхэм Н., Сильверман Э.К., Хейуорд С., Моррис А.П. , Баттерворт А.С., Скотт Р.А., Уолтерс Р.Г., Мейерс Д.А., Чо М.Х., Страчан Д.П., Холл И.П., Тобин М.Д., Уэйн Л.В.Исправление автора: новые генетические сигналы для функции легких выявляют пути и ассоциации хронической обструктивной болезни легких у разных предков. Нат Жене. 2019 июнь; 51(6):1067. PMID: 31110354.
Shrine N, Guyatt AL, Erzurumluoglu AM, Jackson VE, Hobbs BD, Melbourne CA, Batini C, Fawcett KA, Song K, Sakornsakolpat P, Li X, Боксалл Р., Рив Н.Ф., Обейдат М., Чжао Дж. Х., Вильшер М., Вайс С., Кентисту К. А., Кук Дж. П., Сун Б. Б., Чжоу Дж., Хуэй Дж., Карраш С., Имбоден М., Харрис С. Э., Мартен Дж., Энрот С., Керр С.М., Суракка И., Витарт В., Лехтимяки Т., Аллен Р.Дж., Бакке П.С., Бити Т.Х., Бликер Э.Р., Боссе И., Брандсма К.А., Чен З., Крапо Д.Д., Данеш Дж., ДеМео Д.Л., Дадбридж Ф., Эверт Р., Гигер С. , Гулсвик А., Ханселл А.Л., Хао К., Хоффман Дж. Д., Хокансон Дж.Е., Хомут Г., Джоши П.К., Жубер П., Лангенберг С., Ли Х, Ли Л., Лин К., Линд Л., Локанторе Н., Луан Дж., Махаджан А., Маранвиль JC, Мюррей А., Никл Д.С., Пакер Р., Паркер М.М., Пейнтон М.Л., Портеус Д.Дж., Прокопенко Д., Цяо Д., Равал Р., Рунц Х., Сайерс И., Син Д.Д., Смит Б.Х., Солер Артигас М., Воробей Д., Тал- Сингер Р., Тиммерс PRHJ, Ван ден Берге М., Уиттакер Дж.С., Вудрафф П.Г., Йергес-Армстронг Л.М., Троянская О.Г., Райтак Ари О.Т., Кяхёнен М., Полашек О., Гилленстен У., Рудан И., Дири И.Дж., Пробст-Хенш Н.М., Шульц Х., Джеймс А.Л., Уилсон Дж.Ф., Стуббе Б., Зеггини Э., Ярвелин М.Р., Уэрхэм Н., Сильверман Э.К., Хейворд С., Моррис А.П., Баттерворт А.С., Скотт Р.А., Уолтерс Р.Г., Мейерс Д.А., Чо М.Х., Страчан Д.П., Холл И.П., Тобин М.Д., Уэйн Л.В.Новые генетические сигналы для функции легких подчеркивают пути и ассоциации хронической обструктивной болезни легких у разных предков. Нат Жене. 2019 03; 51(3):481-493. PMID: 30804560.
Д., Хокансон Дж.Е., Хомут Г., Джоши П.К., Жубер П., Лангенберг С., Ли Х, Ли Л., Лин К., Линд Л., Локанторе Н., Луан Дж., Махаджан А., Маранвиль JC, Мюррей А., Никл Д.С., Пакер Р., Паркер М.М., Пейнтон М.Л., Портеус Д.Дж., Прокопенко Д., Цяо Д., Равал Р., Рунц Х., Сайерс И., Син Д.Д., Смит Б.Х., Солер Артигас М., Воробей Д., Тал- Сингер Р., Тиммерс PRHJ, Ван ден Берге М., Уиттакер Дж.С., Вудрафф П.Г., Йергес-Армстронг Л.М., Троянская О.Г., Райтак Ари О.Т., Кяхёнен М., Полашек О., Гилленстен У., Рудан И., Дири И.Дж., Пробст-Хенш Н.М., Шульц Х., Джеймс А.Л., Уилсон Дж.Ф., Стуббе Б., Зеггини Э., Ярвелин М.Р., Уэрхэм Н., Сильверман Э.К., Хейворд С., Моррис А.П., Баттерворт А.С., Скотт Р.А., Уолтерс Р.Г., Мейерс Д.А., Чо М.Х., Страчан Д.П., Холл И.П., Тобин М.Д., Уэйн Л.В.Новые генетические сигналы для функции легких подчеркивают пути и ассоциации хронической обструктивной болезни легких у разных предков. Нат Жене. 2019 03; 51(3):481-493. PMID: 30804560.
 02.2019
02.2019Сакорнсаколпат П., Прокопенко Д., Ламонтань М., Рив Н.Ф., Гайатт А.Л., Джексон В.Е., Храм Н., Цяо Д., Барц Т.М., Ким Д.К., Ли MK, Latourelle JC, Li X, Morrow JD, Obeidat M, Wyss AB, Bakke P, Barr RG, Beaty TH, Belinsky SA, Brusselle GG, Crapo JD, de Jong K, DeMeo DL, Fingerlin TE, Gharib SA, Gulsvik A , Холл И.П., Хокансон Дж.Э., Ким В.Дж., Ломас Д.А., Лондон С.Дж., Мейерс Д.А., О'Коннор Г.Т., Реннард С.И., Шварц Д.А., Сливински П., Воробей Д., Страчан Д.П., Тал-Зингер Р., Тесфаигзи Ю., Вестбо Дж., Вонк Дж. М., Йим Дж. Дж., Чжоу Х, Боссе Ю., Маничайкул А., Лахусс Л., Сильверман Э. К., Боезен Х. М., Уэйн Л. В., Тобин М. Д., Хоббс Б. Д., Чо М. Х.Генетический ландшафт хронической обструктивной болезни легких выявляет гетерогенные ассоциации клеточного типа и фенотипа. Нат Жене. 2019 03; 51(3):494-505. PMID: 30804561.
Читайте в: PubMedМассы мошонки | Воробей
Большинство новообразований мошонки требуют минимально инвазивного лечения или вообще не требуют лечения, но некоторые требуют медикаментозного лечения или более серьезных процедур.
Инфекции
Образования мошонки, вызванные бактериальной инфекцией, как это обычно бывает при эпидидимите, лечат антибиотиками.Вирусные инфекции, вызывающие эпидидимит или орхит, обычно лечат отдыхом, льдом и обезболивающими препаратами.
Незлокачественные (доброкачественные) образования мошонки
Доброкачественные образования мошонки можно не лечить или удалять хирургическим путем, восстанавливать или дренировать. Эти решения о лечении зависят от таких факторов, как масса мошонки:
- Вызывает дискомфорт или боль
- Способствует или увеличивает риск бесплодия
- Заражение
Рак яичка
Специалист по лечению рака (онколог) порекомендует лечение в зависимости от того, изолирован ли рак в яичке или распространился на другие ткани организма.Ваш возраст и общее состояние здоровья также являются факторами при выборе вариантов лечения рака яичка.
- Радикальная паховая орхиэктомия. Это основной метод лечения рака яичек.
 Это хирургическая процедура по удалению пораженного яичка и семенного канатика через разрез в паху. Лимфатические узлы в брюшной полости также могут быть удалены, если на них распространился рак.
Это хирургическая процедура по удалению пораженного яичка и семенного канатика через разрез в паху. Лимфатические узлы в брюшной полости также могут быть удалены, если на них распространился рак. - Химиотерапия. Это медикаментозное лечение, в котором используются мощные химические вещества для уничтожения раковых клеток.
 Это хирургическая процедура по удалению пораженного яичка и семенного канатика через разрез в паху. Лимфатические узлы в брюшной полости также могут быть удалены, если на них распространился рак.
Это хирургическая процедура по удалению пораженного яичка и семенного канатика через разрез в паху. Лимфатические узлы в брюшной полости также могут быть удалены, если на них распространился рак.В некоторых случаях также может быть использована лучевая терапия. Этот тип терапии использует высокие дозы рентгеновских лучей или другого высокоэнергетического излучения для уничтожения раковых клеток, которые могут остаться после удаления пораженного яичка.
Большинство случаев рака яичка можно вылечить, но необходимо последующее наблюдение для выявления возможных рецидивов.
Образ жизни и домашние средства
Самостоятельное обследование яичка может помочь вам обнаружить образование в мошонке на ранней стадии, что позволит вам получить своевременную медицинскую помощь.Если вы будете проводить этот осмотр регулярно, вы поймете, что такое «нормальное» ощущение, и будете лучше подготовлены к обнаружению аномалий. Чтобы выполнить самообследование яичка, выполните следующие действия:
Чтобы выполнить самообследование яичка, выполните следующие действия:
- Осматривайте свои яички один раз в месяц, особенно если у вас был рак яичек или у вас есть семейная история рака яичек.
- Выполняйте обследование после принятия теплой ванны или душа. Тепло от воды расслабляет мошонку, облегчая проверку.
- Встаньте перед зеркалом.Ищите припухлость на коже мошонки.
- Накройте мошонку одной рукой, чтобы проверить, отличается ли она от нормальной.
- Осматривайте по одному яичку обеими руками. Поместите указательный и средний пальцы под яичко; поместите большие пальцы сверху.
- Аккуратно покатайте яичко между большими и указательными пальцами, чтобы нащупать уплотнения. Яички обычно гладкие, овальной формы и несколько твердые. Это нормально, если одно яичко немного больше другого.
- Ощупайте мягкую структуру в форме запятой, которая проходит выше и позади яичка (придатка яичка), чтобы проверить, нет ли припухлости.

Если вы обнаружите припухлость или другую аномалию, как можно скорее обратитесь к врачу.
Подготовка к встрече
Если вы чувствуете боль, обратитесь за неотложной помощью. Если вы обнаружите новообразование в мошонке, вы, вероятно, начнете с посещения семейного врача. Вас могут направить к специалисту по заболеваниям мочевыводящих путей и мужских половых органов (урологу).
Подготовка к приему вас или вашего ребенка к врачу или урологу поможет вам максимально эффективно использовать время, проведенное с врачом.
Что вы можете сделать
Запишите информацию, чтобы поделиться с врачом, в том числе:
- Симптомы , которые вы испытываете, включая любые, которые могут показаться не связанными с опухолью мошонки
- Ключевая личная информация, включая серьезные стрессы или недавние изменения в жизни
- Лекарства, витамины и добавки, которые вы принимаете
- Семейный анамнез рака яичек или других заболеваний мошонки
- Личный анамнез, включая предшествующие новообразования мошонки, неопустившееся яичко или врожденные дефекты, связанные с гениталиями
- Вопросы к врачу
Вопросы о новообразованиях мошонки могут включать:
- Какие анализы мне понадобятся?
- Сколько времени потребуется, чтобы получить результаты теста?
- Если опухоль мошонки раковая (злокачественная), каковы следующие шаги?
- Если опухоль мошонки не раковая, нужно ли мне лечение?
- Могу ли я взять с собой брошюры или другие печатные материалы? Какие сайты вы рекомендуете?
Не стесняйтесь задавать любые другие вопросы.
Чего ожидать от вашего врача
Ваш врач, вероятно, задаст вам ряд вопросов, в том числе:
- Когда вы обнаружили уплотнение или другие симптомы, связанные с опухолью мошонки?
- У вас есть или были боли в мошонке или рядом с ней?
- Была ли у вас лихорадка, кровь или гной в моче?
- У вас была недавняя травма паха?
- Что-нибудь, например обезболивающее, улучшает ваши симптомы?
- Ухудшает ли что-либо симптомы, например, упражнения или нагрузки, вызывающие напряжение в паху?
- Было ли у вас неопущение или втяжение яичка, которое было исправлено хирургическим путем?
- Были ли у Вас когда-либо инфекции, передающиеся половым путем?
- У вас есть несколько сексуальных партнеров или новый сексуальный партнер?
House Sparrow (EwA Guide to the Birds of the Fells (Массачусетс, США)) · iNaturalist
Резюме
5 Домашний воробей ( Passer domesticus ) — птица семейства воробьиных воробьиных, обитающая в большинстве частей мира. Небольшая птица имеет типичную длину 16 см (6,3 дюйма) и массу 24–39,5 г (0,85–1,39 унции). Самки и молодые птицы окрашены в бледно-коричневый и серый цвета, а самцы имеют более яркие черные, белые и коричневые отметины. Один из примерно 25 видов рода Passer , домашний воробей является родным для большей части Европы, Средиземноморского бассейна,...
Небольшая птица имеет типичную длину 16 см (6,3 дюйма) и массу 24–39,5 г (0,85–1,39 унции). Самки и молодые птицы окрашены в бледно-коричневый и серый цвета, а самцы имеют более яркие черные, белые и коричневые отметины. Один из примерно 25 видов рода Passer , домашний воробей является родным для большей части Европы, Средиземноморского бассейна,...
Биология таксона
6Домашний воробей ( Passer domesticus ) произрастает в Евразии и на северной окраине африканского континента, но в настоящее время встречается в большинстве регионов мира, где живут люди.Этот вид в основном связан с людьми, живущими вокруг зданий в различных условиях, от изолированных сельских ферм до крупных городских центров, хотя плотность населения и успех размножения обычно выше в пригородной среде, чем в городах или сельской местности.
Домовые воробьи питаются в основном вегетарианской пищей, питаясь в основном семенами сорняков и трав или отходами зерна, а также почками, ягодами и различными отходами от людей. Летом животный материал может составлять до 10% рациона, и, как оппортунисты, домовые воробьи могут питаться маленькими лягушками.моллюсков и ракообразных, если таковые имеются. Птенцов в первые дни после вылупления кормят главным образом насекомыми.
Летом животный материал может составлять до 10% рациона, и, как оппортунисты, домовые воробьи могут питаться маленькими лягушками.моллюсков и ракообразных, если таковые имеются. Птенцов в первые дни после вылупления кормят главным образом насекомыми.
Домовые воробьи, как правило, собираются в стаи, часто довольно большие вне сезона размножения. Гнездятся в основном разрозненными колониями от 10 до 20 пар. Строительство гнезда инициирует неспаренный самец, но после образования пары ему помогает его самка. Гнезда обычно строят в искусственных или естественных полостях или расщелинах. Эти птицы удивительно католичны в выборе места для гнезда: сообщается о гнездах с движущихся машин и даже с глубины 640 м под землей в угольной шахте в Англии.Размер кладки обычно составляет от 3 до 6 яиц. Насиживание (от 10 до 14 дней) осуществляется обоими родителями. Оба родителя также кормят птенцов, которые покидают гнездо примерно через две недели после вылупления. Обычно в год производят две-три кладки.
Домовые воробьи не мигрируют на большей части своего естественного и интродуцированного ареала. Этот вид является одним из самых многочисленных птиц в мире: общая европейская популяция в 1980-х и 1990-х годах, вероятно, превышала 50 000 000 размножающихся пар, а общая мировая численность оценивается примерно в 500 000 000 пар.Однако в последние годы сообщалось о значительном сокращении популяции в некоторых частях местного и интродуцированного ареала (хотя расширение ареала в других местах продолжалось).
(Кауфман, 1996; Саммерс-Смит, 2009 и ссылки в них)
Источники и Кредиты
- (c) Билл МакИндевар, некоторые права защищены (CC BY-NC), https://www.inaturalist.org/photos/64986749
- (c) Билл МакИндевар, некоторые права защищены (CC BY-NC), https://www.inaturalist.org/photos/64986758
- (c) Лип Ки Яп, некоторые права защищены (CC BY-SA),
- (c) Кейт Маршалл, некоторые права защищены (CC BY-NC-SA),
http://farm3. static.flickr.com/2729/4143585089_2a01b5bfbf.jpg
- (c) Википедия, некоторые права защищены (CC BY-SA), http://en.wikipedia.org/wiki/Passer_domesticus
- Адаптировано Клэр О'Нил из произведения (c) Unknown, некоторые права защищены (CC BY-NC), http://eol.org/data_objects/17428264

 static.flickr.com/2729/4143585089_2a01b5bfbf.jpg
static.flickr.com/2729/4143585089_2a01b5bfbf.jpgПодробнее
Missa brevis in C K220 'Воробьиная месса' Вольфганга Амадея Моцарта » Ноты для оркестра (партитура)
Missa brevis in C K220 'Воробьиная месса' Вольфганга Амадея Моцарта » Ноты для оркестра (партитура)- Ноты для инструментов
- Хор, Вокал
- Ансамбль
- Теория, Книги, Аксессуары
- Загрузки
- Распродажа
- Информация
- О нотах. ком
Контакт
- О нотах.
- Магазин
 ком
комграниц | Интегративные физиологические реакции на острую дегидратацию у красношейного воробья: метаболические, ферментативные и окислительные признаки
Введение
Прогнозируется, что учащение засух и волн тепла окажет сильное влияние на фенологию, поведение и физиологию растений и животных (Walther et al. , 2002; Олбрайт и др., 2017). Хотя прогнозы показывают, что все птицы во всем мире будут затронуты глобальным потеплением и экстремальными климатическими явлениями, виды, обитающие в субтропических и умеренных пустынях, будут непропорционально затронуты изменением климата, поскольку величина и, возможно, скорость изменений в этих регионах, вероятно, будут больше и быстрее, чем в других биомах (McKechnie, Wolf, 2010; Albright et al., 2017; Iknayan, Beissinger, 2018). Это особенно актуально для воробьиных, потому что общее поведение (т.g., дневные), эволюционные (например, небольшой размер тела) и физиологические (например, высокая удельная скорость метаболизма, рассеивание тепла через тяжелое дыхание) черты этой группы делают ее особенно восприимчивой к повышению температуры и засушливости (Йом-Тов). , 2001; Гарднер и др., 2011; Олбрайт и др., 2017). Чтобы лучше оценить, насколько быстрые изменения окружающей среды угрожают выживанию видов, феноменологические данные следует объединить с лабораторными экспериментами, чтобы определить механизмы, контролирующие реакцию животных на условия окружающей среды (Khaliq et al.
, 2002; Олбрайт и др., 2017). Хотя прогнозы показывают, что все птицы во всем мире будут затронуты глобальным потеплением и экстремальными климатическими явлениями, виды, обитающие в субтропических и умеренных пустынях, будут непропорционально затронуты изменением климата, поскольку величина и, возможно, скорость изменений в этих регионах, вероятно, будут больше и быстрее, чем в других биомах (McKechnie, Wolf, 2010; Albright et al., 2017; Iknayan, Beissinger, 2018). Это особенно актуально для воробьиных, потому что общее поведение (т.g., дневные), эволюционные (например, небольшой размер тела) и физиологические (например, высокая удельная скорость метаболизма, рассеивание тепла через тяжелое дыхание) черты этой группы делают ее особенно восприимчивой к повышению температуры и засушливости (Йом-Тов). , 2001; Гарднер и др., 2011; Олбрайт и др., 2017). Чтобы лучше оценить, насколько быстрые изменения окружающей среды угрожают выживанию видов, феноменологические данные следует объединить с лабораторными экспериментами, чтобы определить механизмы, контролирующие реакцию животных на условия окружающей среды (Khaliq et al. , 2014).
, 2014).
Поддержание осмотического баланса является сложной задачей, когда птицы питаются соленой пищей или живут в засушливых местообитаниях с дефицитом пресной воды (Sabat and Martinez del Rio, 2002; Sabat et al., 2009; Smit and McKechnie, 2015). Водный баланс птиц является функцией взаимодействия между доступностью воды в окружающей среде, физиологическими и поведенческими механизмами, которые птицы используют для сокращения потерь воды, и производством метаболической воды, которая напрямую связана со скоростью метаболизма (Gerson and Guglielmo, 2011; Albright et al. др., 2017; Сабат и др., 2021). Некоторые виды птиц реагируют на условия окружающей среды, которые усиливают осморегуляцию, увеличивая скорость расхода энергии, включая скорость основного обмена (BMR). Этот ответ был связан с увеличением гиперосмотической экскреции мочи, что, в свою очередь, включает как затраты энергии на осморегуляцию per se (McWhorter et al., 2004), так и долгосрочный эффект увеличения массы и интенсивности метаболизма тканей. таких как почки и сердце (Daan et al., 1990; Гутьеррес и др., 2011 г.; Пенья-Вильялобос и др., 2013 г.; Сабат и др., 2017). Тем не менее, влияние обезвоживания на скорость метаболизма у птиц дало противоречивые результаты (Cade et al., 1965; Gerson and Guglielmo, 2011).
таких как почки и сердце (Daan et al., 1990; Гутьеррес и др., 2011 г.; Пенья-Вильялобос и др., 2013 г.; Сабат и др., 2017). Тем не менее, влияние обезвоживания на скорость метаболизма у птиц дало противоречивые результаты (Cade et al., 1965; Gerson and Guglielmo, 2011).
Еще одной потенциальной причиной обезвоживания может быть окислительный стресс, возникающий в результате поддержания высокого уровня метаболизма. Митохондрии являются основным источником клеточных активных форм кислорода (АФК), связанных с продукцией анионов супероксида в качестве побочных продуктов цепи переноса электронов и других окислительно-восстановительных реакций (Srinivasan and Avadhani, 2012).В нескольких исследованиях сообщалось, что увеличение скорости метаболизма приводит к более высокой продукции АФК, что, следовательно, стимулирует энергоемкую антиоксидантную защиту (Commoner et al., 1954; Cohen et al., 2008; Tumminello and Fuller-Espie, 2013). Более того, некоторые исследования показали, что осмотический стресс влияет на несколько физиологических признаков у птиц, включая сдвиги в тканеспецифичной скорости метаболизма и окислительного статуса (Tieleman et al. , 2003; Tapia-Monsalve et al., 2018; Sabat et al., 2019). ).В поддержку этого механизма недавнее исследование показало, что исходные уровни кортикостерона были выше у домашних воробьев, лишенных воды (Brischoux et al., 2020), и было показано, что глюкокортикоиды нарушают клеточный окислительный гомеостаз (Costantini et al., 2011; Queisser). и др., 2011). Кроме того, недавние данные указывают на то, что обезвоживание, вызванное высокими температурами окружающей среды, влияет на клеточный метаболизм у млекопитающих, что в конечном итоге приводит к усилению антиоксидантной реакции и/или окислительному повреждению (Jacobs et al., 2020). Остается неизвестным, могут ли состояние гидратации и изменения осмотических условий вызывать окислительное повреждение у птиц. Мы предсказали, что потенциальной дополнительной стоимостью обезвоживания является увеличение производства АФК, что приводит к кумулятивному повреждению биомолекул, Costantini et al. (2010).
, 2003; Tapia-Monsalve et al., 2018; Sabat et al., 2019). ).В поддержку этого механизма недавнее исследование показало, что исходные уровни кортикостерона были выше у домашних воробьев, лишенных воды (Brischoux et al., 2020), и было показано, что глюкокортикоиды нарушают клеточный окислительный гомеостаз (Costantini et al., 2011; Queisser). и др., 2011). Кроме того, недавние данные указывают на то, что обезвоживание, вызванное высокими температурами окружающей среды, влияет на клеточный метаболизм у млекопитающих, что в конечном итоге приводит к усилению антиоксидантной реакции и/или окислительному повреждению (Jacobs et al., 2020). Остается неизвестным, могут ли состояние гидратации и изменения осмотических условий вызывать окислительное повреждение у птиц. Мы предсказали, что потенциальной дополнительной стоимостью обезвоживания является увеличение производства АФК, что приводит к кумулятивному повреждению биомолекул, Costantini et al. (2010).
Клетки могут сокращаться в ответ на повышение осмоляльности плазмы крови при водном стрессе, а эритроциты должны поддерживать свой объем за счет энергоемкой ионной регуляции (Hoffmann et al. , 2009; Герсон и Гульельмо, 2011). У птиц в красных кровяных тельцах (эритроцитах) есть функционирующие митохондрии, содержащие специфические ферменты, которые немедленно обеспечивают энергией транспортирующие ионы белки (Moritz et al., 1997; Stier et al., 2013). Недавние исследования документально подтвердили существование значительных межиндивидуальных различий в расходе энергии митохондриями эритроцитов у птиц. В частности, изменение митохондриального метаболизма связано с активностью метаболических ферментов цитратсинтазы (CS) и цитохром-С-оксидазы (ЦОГ) в скелетных мышцах (Stier et al., 2015, 2017). Эти два фермента отвечают за выработку энергии (АТФ), поэтому активность этих ферментов коррелирует со скоростью потребления энергии всем телом птиц (Peña-Villalobos et al., 2014; Gutiérrez et al., 2019). Таким образом, возможно, что метаболизм митохондрий посредством экспрессии ферментов, участвующих в расходовании энергии на тканевом и периферическом уровне, коррелирует с физиологическими изменениями, происходящими у птиц, подвергающихся водному стрессу.
, 2009; Герсон и Гульельмо, 2011). У птиц в красных кровяных тельцах (эритроцитах) есть функционирующие митохондрии, содержащие специфические ферменты, которые немедленно обеспечивают энергией транспортирующие ионы белки (Moritz et al., 1997; Stier et al., 2013). Недавние исследования документально подтвердили существование значительных межиндивидуальных различий в расходе энергии митохондриями эритроцитов у птиц. В частности, изменение митохондриального метаболизма связано с активностью метаболических ферментов цитратсинтазы (CS) и цитохром-С-оксидазы (ЦОГ) в скелетных мышцах (Stier et al., 2015, 2017). Эти два фермента отвечают за выработку энергии (АТФ), поэтому активность этих ферментов коррелирует со скоростью потребления энергии всем телом птиц (Peña-Villalobos et al., 2014; Gutiérrez et al., 2019). Таким образом, возможно, что метаболизм митохондрий посредством экспрессии ферментов, участвующих в расходовании энергии на тканевом и периферическом уровне, коррелирует с физиологическими изменениями, происходящими у птиц, подвергающихся водному стрессу.
Здесь мы интегрировали измерение энергетического метаболизма по крови (например,g., CS и COX) с окислительным статусом (например, перекисное окисление липидов и баланс глутатиона) и BMR для изучения осморегуляторных приспособлений к обезвоживанию у рыжешейных воробьев в неволе ( Zonotrichia capensis ), обычных воробьиных птиц, обитающих на юге Южной Америки. В частности, мы предполагаем, что птицы, подвергшиеся ограничению воды в течение 2 недель, будут демонстрировать более высокую осмоляльность плазмы и мочи, активность митохондриальных ферментов и скорость метаболизма, чем гидратированные (контрольные) птицы.Мы также прогнозируем, что у птиц с ограничением воды (WR) должно быть более высокое соотношение окисленного (GSSG) к восстановленному (GSH) глутатиона; более высокое перекисное окисление липидов (концентрация MDA) как показатель окислительного повреждения, более высокие концентрации пероксидов водорода (H 2 O 2 ) и повышенная общая антиоксидантная способность (TAC).
Материалы и методы
Лаборатория акклиматизации
Всего в ноябре 2018 г. было отловлено 18 особей Z. capensis (Passeriformes: Emberizidae).Птиц в неволе в течение 2 недель содержали при постоянной температуре 25 ± 2 ° С в пределах их термонейтральной зоны (ТНЗ; Sabat et al., 2019) и содержали индивидуально в клетках из пластиковой сетки (35 см × 35 см × 35 см). ) при фотопериоде 12L:12D. После отлова каждой птице была назначена одна из двух обработок: контрольная (CT) группа содержалась с водой и семенами ad libitum и группа WR с семенами ad libitum , которым в течение первой недели разрешалось пить свежую воду. каждые 24 часа, а вторую неделю каждые 48 часов.Людей взвешивали на весах (±0,001 г) в начале и в конце каждой недели лечения. Птицы были отловлены с разрешениями, выданными Servicio Agricola Ganadero (№ 10192/2019). Все протоколы были одобрены институциональным комитетом по уходу за животными Чилийского университета, где проводились эксперименты.
Скорость основного обмена и общая потеря воды при испарении
Сразу после каждой обработки мы измеряли BMR (мл O 2 ч –1 ) и общую потерю воды при испарении (TEWL) у птиц после абсорбции (натощак в течение 4 часов) в течение ночи между 21:00 и 07:00 с использованием стандартных методов проточной респирометрии.Измерения респирометрии проводились на трех птицах за ночь и располагались в шахматном порядке таким образом, чтобы начало экспериментов по ограничению воды начиналось последовательно. Потребление кислорода измеряли с помощью трех компьютеризированных респирометрических систем с открытым потоком (две FMS, одна FOXBOX, Sable Systems, Хендерсон, Невада, США), откалиброванных с использованием известной смеси кислорода (20%) и азота (80%), которые были сертифицированы хроматографии (INDURA, Чили), а затем помещают в шкаф с регулируемой температурой (Sable Systems, Henderson, Nevada) при постоянной температуре окружающей среды 30±0.5 ° С, что находится в пределах ТНЗ для этого вида (Sabat et al. , 2019). В метаболическую камеру подавался сухой воздух в количестве 500 мл мин –1 через трубку Bev-A-Line (Thermoplastic Processes Inc) от регулятора массового расхода. Выходящий воздух проходил через гигрометр, регистрирующий давление паров (кПа), а затем через колонки с баралимом для поглощения СО 2 перед прохождением через анализатор O 2 . Поскольку CO 2 очищался перед подачей в анализатор O 2 , потребление кислорода рассчитывалось как (Withers, 1977): V . O2 = [FR × 60 × (Fi O 2 –Fe O 2 )]/(1–Fi O 2 )], где FR – скорость потока в мл/мин, Fi и Fe – фракционные концентрации O 2 на входе и выходе из метаболической камеры соответственно. TEWL рассчитывали как TEWL = [(Ve ρ из – Vi ρ из )], где TEWL выражен в мг/мл. ρвх и ρвых – абсолютная влажность в кг/м 3 воздуха на входе и воздухе на выходе соответственно, Ve – расход воздуха, поступающего в камеру, по данным регулятора массового расхода, оснащенного FMS и FOXBOX ( 500 мл мин –1 ), Ve – расход выходящего воздуха.
, 2019). В метаболическую камеру подавался сухой воздух в количестве 500 мл мин –1 через трубку Bev-A-Line (Thermoplastic Processes Inc) от регулятора массового расхода. Выходящий воздух проходил через гигрометр, регистрирующий давление паров (кПа), а затем через колонки с баралимом для поглощения СО 2 перед прохождением через анализатор O 2 . Поскольку CO 2 очищался перед подачей в анализатор O 2 , потребление кислорода рассчитывалось как (Withers, 1977): V . O2 = [FR × 60 × (Fi O 2 –Fe O 2 )]/(1–Fi O 2 )], где FR – скорость потока в мл/мин, Fi и Fe – фракционные концентрации O 2 на входе и выходе из метаболической камеры соответственно. TEWL рассчитывали как TEWL = [(Ve ρ из – Vi ρ из )], где TEWL выражен в мг/мл. ρвх и ρвых – абсолютная влажность в кг/м 3 воздуха на входе и воздухе на выходе соответственно, Ve – расход воздуха, поступающего в камеру, по данным регулятора массового расхода, оснащенного FMS и FOXBOX ( 500 мл мин –1 ), Ve – расход выходящего воздуха. Ve рассчитывали согласно (Tieleman et al., 2003) как Ve = Vi –[V . O 2 (1–RQ)] + VH 2 O Vi и V . O 2 (мл мин –1 ). Мы использовали дыхательный коэффициент (RQ) равный 0,71, что предполагает, что голодающие воробьи в основном полагаются на накопленные липиды для метаболизма (King and Farner, 1961; Walsberg and Wolf, 1995). Абсолютная влажность рассчитывалась как ρ = P/(T × Rw), где P — давление водяного пара в воздухе в Паскалях, T — температура гигрометра точки росы в Кельвинах, а Rw — газовая постоянная для водяного пара. [461.5 Дж/кг × Кельвин, (Lide, 2001)]. ρ в определяли, используя среднее значение давления паров воздуха, поступающего в пустую камеру в течение 15-минутного базового периода до и после каждого эксперимента. Перед метаболическими измерениями измеряли массу тела с помощью электронных весов (±0,1 г), а клоакальную температуру тела (Tb) регистрировали тонкой медно-константановой термопарой Коула-Палмера, присоединенной к термометру Digisense (модель 92800-15) в течение минуты.
Ve рассчитывали согласно (Tieleman et al., 2003) как Ve = Vi –[V . O 2 (1–RQ)] + VH 2 O Vi и V . O 2 (мл мин –1 ). Мы использовали дыхательный коэффициент (RQ) равный 0,71, что предполагает, что голодающие воробьи в основном полагаются на накопленные липиды для метаболизма (King and Farner, 1961; Walsberg and Wolf, 1995). Абсолютная влажность рассчитывалась как ρ = P/(T × Rw), где P — давление водяного пара в воздухе в Паскалях, T — температура гигрометра точки росы в Кельвинах, а Rw — газовая постоянная для водяного пара. [461.5 Дж/кг × Кельвин, (Lide, 2001)]. ρ в определяли, используя среднее значение давления паров воздуха, поступающего в пустую камеру в течение 15-минутного базового периода до и после каждого эксперимента. Перед метаболическими измерениями измеряли массу тела с помощью электронных весов (±0,1 г), а клоакальную температуру тела (Tb) регистрировали тонкой медно-константановой термопарой Коула-Палмера, присоединенной к термометру Digisense (модель 92800-15) в течение минуты. после удаления птиц из метаболической камеры.Мы обнаружили, что все животные были эутермичны после метаболических испытаний (Tb > 40 ° C). Считывание данных проводили с помощью программного обеспечения EXPEDATA (Sable Systems, International, NV, США). Чтобы оценить скорость основного обмена и TEWL, мы усреднили концентрации O 2 и давление водяного пара в вытекающем воздушном потоке в течение 20-минутного периода после достижения устойчивого состояния, что обычно происходит через 3 часа у Z. capensis (Maldonado et al. др., 2009). Мы оценили производство метаболической воды (МВП) воробьев, используя эквивалентность 0.567 мл H 2 O на литр потребленного O 2 (Schmidt-Nielsen, 1997). Мы рассчитали соотношение между MWP и потерями воды (MWP/TEWL) за 20-минутный период, в течение которого было достигнуто стационарное состояние.
после удаления птиц из метаболической камеры.Мы обнаружили, что все животные были эутермичны после метаболических испытаний (Tb > 40 ° C). Считывание данных проводили с помощью программного обеспечения EXPEDATA (Sable Systems, International, NV, США). Чтобы оценить скорость основного обмена и TEWL, мы усреднили концентрации O 2 и давление водяного пара в вытекающем воздушном потоке в течение 20-минутного периода после достижения устойчивого состояния, что обычно происходит через 3 часа у Z. capensis (Maldonado et al. др., 2009). Мы оценили производство метаболической воды (МВП) воробьев, используя эквивалентность 0.567 мл H 2 O на литр потребленного O 2 (Schmidt-Nielsen, 1997). Мы рассчитали соотношение между MWP и потерями воды (MWP/TEWL) за 20-минутный период, в течение которого было достигнуто стационарное состояние.
Осмоляльность крови и мочи
Кровь (50–100 мкл) забирали утром (09:00–11:00) перед физиологическими измерениями из плечевой вены с помощью гематокритных пробирок с антикоагулянтом (гепарином или ЭДТА).:max_bytes(150000):strip_icc()/house-sparrow_3-5a0dd9a14e4f7d00367cdd4a.jpg) Образцы крови центрифугировали при 10000 g в течение 5 мин для отделения плазмы от эритроцитов, и обе фракции замораживали при -80 ° С до проведения анализа.Мочу из мочеточника получали путем введения маленькой канюли с закрытым концом в клоаку птицы, которую центрифугировали, а надосадочную жидкость собирали для осмометрического анализа (Wescor 5130B). После метаболических измерений птиц забивали воздействием СО 2 , взвешивали, извлекали органы и замораживали при -80 ° С.
Образцы крови центрифугировали при 10000 g в течение 5 мин для отделения плазмы от эритроцитов, и обе фракции замораживали при -80 ° С до проведения анализа.Мочу из мочеточника получали путем введения маленькой канюли с закрытым концом в клоаку птицы, которую центрифугировали, а надосадочную жидкость собирали для осмометрического анализа (Wescor 5130B). После метаболических измерений птиц забивали воздействием СО 2 , взвешивали, извлекали органы и замораживали при -80 ° С.
Метаболические ферменты
Осадок крови, состоящий из клеточной фракции, состоящей в основном из эритроцитов, оттаивали и переносили в пробирку Falcon, взвешивали на аналитических весах (±0.0001) и гомогенизировали на льду (1:4 мас./об.) в 0,1 М фосфатном буфере с добавлением 2 мМ ЭДТА (рН 7,3). Чтобы получить аналогичные массы изолятов эритроцитов в образцах, вес данного осадка эритроцитов был скорректирован на значение среднего гематокрита всех образцов (т. е. w × [гематокрит образца/средний гематокрит]). Затем образцы обрабатывали ультразвуком с использованием ультразвукового процессора мощностью 130 Вт (VCX 130) на льду 14 раз в 20-секундных циклах с 10-секундным интервалом между циклами. Затем гомогенаты центрифугировали при 14000 об/мин в течение 15 мин при 4° С для получения постмитохондриальной фракции.Супернатант переносили в новую пробирку, чтобы избежать переноса верхнего липидного слоя, присутствующего в гомогенате. Концентрации белка определяли по методу Брэдфорда (1976) с использованием альбумина бычьей сыворотки в качестве стандарта. Мы измерили активность двух митохондриальных ферментов: ЦОГ (КФ 1.9.3.1) и ХС (КФ 4.1.3.7). Активность ЦОГ количественно определяли с помощью спектрофотометрического метода на микропланшетах, в котором активность фермента измеряли в реакционной смеси, содержащей 10 мМ трис-HCl (pH 7), 120 мМ KCl, 250 мМ сахарозы и цитохром с, восстановленный дитиотреитом, в конечном объеме 0 .2 мл. Уменьшение экстинкции при 550 нм контролировали на спектрофотометре Thermo Scientific Multiskan GO UV/VIS при 25 ° C.
е. w × [гематокрит образца/средний гематокрит]). Затем образцы обрабатывали ультразвуком с использованием ультразвукового процессора мощностью 130 Вт (VCX 130) на льду 14 раз в 20-секундных циклах с 10-секундным интервалом между циклами. Затем гомогенаты центрифугировали при 14000 об/мин в течение 15 мин при 4° С для получения постмитохондриальной фракции.Супернатант переносили в новую пробирку, чтобы избежать переноса верхнего липидного слоя, присутствующего в гомогенате. Концентрации белка определяли по методу Брэдфорда (1976) с использованием альбумина бычьей сыворотки в качестве стандарта. Мы измерили активность двух митохондриальных ферментов: ЦОГ (КФ 1.9.3.1) и ХС (КФ 4.1.3.7). Активность ЦОГ количественно определяли с помощью спектрофотометрического метода на микропланшетах, в котором активность фермента измеряли в реакционной смеси, содержащей 10 мМ трис-HCl (pH 7), 120 мМ KCl, 250 мМ сахарозы и цитохром с, восстановленный дитиотреитом, в конечном объеме 0 .2 мл. Уменьшение экстинкции при 550 нм контролировали на спектрофотометре Thermo Scientific Multiskan GO UV/VIS при 25 ° C. Активность ЦОГ рассчитывали с использованием коэффициента экстинкции 21,84 мМ –1 см –1 при 550 нм. Для количественного определения CS среда для ферментативного анализа содержала 10 мМ Трис-HCl (pH 8,0), 10 мМ 5,5'-дитиобис-(2-нитробензойная кислота), 30 мМ ацетил-кофермента А (ацетил-КоА) и 10 мМ щавелевоуксусной кислоты (ОАА). в конечном объеме 0,2 мл; эти реагенты были исключены в контроле.CS катализирует реакцию между ацетил-КоА и ОАА с образованием лимонной кислоты. Увеличение экстинкции при 412 нм измеряли в Thermo Scientific Multiskan GO при 25 ° C. Активность CS рассчитывали с использованием коэффициента экстинкции 13,6 мМ –1 см –1 при 412 нм (Peña-Villalobos et al. др., 2014).
Активность ЦОГ рассчитывали с использованием коэффициента экстинкции 21,84 мМ –1 см –1 при 550 нм. Для количественного определения CS среда для ферментативного анализа содержала 10 мМ Трис-HCl (pH 8,0), 10 мМ 5,5'-дитиобис-(2-нитробензойная кислота), 30 мМ ацетил-кофермента А (ацетил-КоА) и 10 мМ щавелевоуксусной кислоты (ОАА). в конечном объеме 0,2 мл; эти реагенты были исключены в контроле.CS катализирует реакцию между ацетил-КоА и ОАА с образованием лимонной кислоты. Увеличение экстинкции при 412 нм измеряли в Thermo Scientific Multiskan GO при 25 ° C. Активность CS рассчитывали с использованием коэффициента экстинкции 13,6 мМ –1 см –1 при 412 нм (Peña-Villalobos et al. др., 2014).
Окислительный стресс
Ткани размораживали и обрабатывали в соответствии со специальными процедурами для каждого аналитического протокола (см. ниже). Окислительный стресс оценивали путем измерения концентрации восстановленного (GSH) и окисленного (GSSG) глутатиона в грудной клетке и печени. Перекисное окисление липидов (TBARS) измеряли в плазме, печени, сердце, грудной клетке и эритроцитах. Пероксиды водорода (H 2 O 2 ) измеряли в плазме, печени, сердце и грудной клетке. Наконец, мы измерили ТАС в плазме, сердце и грудной клетке.
Перекисное окисление липидов (TBARS) измеряли в плазме, печени, сердце, грудной клетке и эритроцитах. Пероксиды водорода (H 2 O 2 ) измеряли в плазме, печени, сердце и грудной клетке. Наконец, мы измерили ТАС в плазме, сердце и грудной клетке.
Концентрацию восстановленного (GSH) и окисленного (GSSG) глутатиона измеряли в кислых супернатантах, которые были получены путем обработки постмитохондриальной фракции (200 мкл) 200 мкл холодной 0,6% (масса/объем) сульфосалициловой кислоты, и центрифугированный (10.000 × г , 5 мин, 4 ° В) после выдерживания образцов в течение 5 мин на льду. Определение концентраций GSH и GSSG проводилось колориметрическим методом Rahman et al. (2006), адаптированный для формата микропланшетов. Перекисное окисление липидов измеряли в плазме, печени, сердце, грудной клетке и эритроцитах (EF) с помощью коммерческого набора Oxiselect, STA-330 Cell Biolabs; (Гутьеррес и др., 2019). Анализ оценивает соединение между аддуктом. Поглощение измеряли на спектрофотометре Thermo Scientific Multiskan GO UV/VIS при 25 ° C.Мы измерили пероксиды водорода (H 2 O 2 ) в плазме как показатель прооксидантного статуса (Costantini, 2014) с использованием коммерческого набора (MAK311, Sigma Aldrich). В анализе колориметрически оценивали окисление Fe +2 до Fe +3 пероксидами, присутствующими в образце, при 585 нм. Мы также измерили антиоксидантную способность с поправкой на титрационный микропланшет (Ribeiro et al., 2011). Вкратце, анализ измеряет восстановление комплекса Cu(II)-неокупроин до комплекса Cu(I)-неокупроин антиоксидантами, присутствующими в образце, анализ измеряется спектрофотометрически при 450 нм, и значение образца сравнивается с известным Trolox. стандартная кривая.
Поглощение измеряли на спектрофотометре Thermo Scientific Multiskan GO UV/VIS при 25 ° C.Мы измерили пероксиды водорода (H 2 O 2 ) в плазме как показатель прооксидантного статуса (Costantini, 2014) с использованием коммерческого набора (MAK311, Sigma Aldrich). В анализе колориметрически оценивали окисление Fe +2 до Fe +3 пероксидами, присутствующими в образце, при 585 нм. Мы также измерили антиоксидантную способность с поправкой на титрационный микропланшет (Ribeiro et al., 2011). Вкратце, анализ измеряет восстановление комплекса Cu(II)-неокупроин до комплекса Cu(I)-неокупроин антиоксидантами, присутствующими в образце, анализ измеряется спектрофотометрически при 450 нм, и значение образца сравнивается с известным Trolox. стандартная кривая.
Количественный магнитно-резонансный анализ
Для оценки влияния обезвоживания на состав тела мы повторили эксперимент по ограничению воды на второй группе Z. capensis ( n = 11), отловленных в том же месте и в тех же экспериментальных условиях. По логистическим причинам мы не могли проанализировать состав тела птиц в начальном эксперименте. В конце этого второго эксперимента по акклиматизации, в ночь после последнего периода обезвоживания (т.д., день 14) птиц перевезли в Университет Чили в Вальдивии (около 850 км от Сантьяго, 10 часов пути) и поместили в аппарат количественного магнитного резонанса (QMR) EchoMRI 500 (Хьюстон, Техас, США). прибор для непосредственного измерения граммов мышечной массы, жировой массы, а также общей и свободной воды в организме. Этот инструмент был апробирован на диких животных (Riley et al., 2016; Kraft et al., 2019; Eastick et al., 2020) и специально использовался для измерения изменений нежировой и жировой массы во время миграции у птиц (Guglielmo et al. др., 2011; Келси и Байрлейн, 2019 г.). Для каждого измерения животное помещали в акриловый цилиндр (диаметром 5 см, длиной 60 см) и иммобилизовали с помощью поршня с липучкой, помещенного внутрь цилиндра. Затем цилиндр помещали в магнитно-резонансный модуль, который был запрограммирован на три сканирования на животное, достигаемые примерно за 1,5 мин.
По логистическим причинам мы не могли проанализировать состав тела птиц в начальном эксперименте. В конце этого второго эксперимента по акклиматизации, в ночь после последнего периода обезвоживания (т.д., день 14) птиц перевезли в Университет Чили в Вальдивии (около 850 км от Сантьяго, 10 часов пути) и поместили в аппарат количественного магнитного резонанса (QMR) EchoMRI 500 (Хьюстон, Техас, США). прибор для непосредственного измерения граммов мышечной массы, жировой массы, а также общей и свободной воды в организме. Этот инструмент был апробирован на диких животных (Riley et al., 2016; Kraft et al., 2019; Eastick et al., 2020) и специально использовался для измерения изменений нежировой и жировой массы во время миграции у птиц (Guglielmo et al. др., 2011; Келси и Байрлейн, 2019 г.). Для каждого измерения животное помещали в акриловый цилиндр (диаметром 5 см, длиной 60 см) и иммобилизовали с помощью поршня с липучкой, помещенного внутрь цилиндра. Затем цилиндр помещали в магнитно-резонансный модуль, который был запрограммирован на три сканирования на животное, достигаемые примерно за 1,5 мин. Если коэффициент вариации этих сканов превышал 6%, обычно из-за движения животного внутри зонда, измерение отбрасывали и повторяли. Мы калибровали QMR ежедневно перед каждой серией измерений в соответствии с рекомендацией производителя, используя образец канолового масла известной массы.Мы рассчитали индекс гидратации для каждой отдельной птицы как: общая вода – свободная вода/нежировая масса.
Если коэффициент вариации этих сканов превышал 6%, обычно из-за движения животного внутри зонда, измерение отбрасывали и повторяли. Мы калибровали QMR ежедневно перед каждой серией измерений в соответствии с рекомендацией производителя, используя образец канолового масла известной массы.Мы рассчитали индекс гидратации для каждой отдельной птицы как: общая вода – свободная вода/нежировая масса.
Статистический анализ
Мы использовали дисперсионный анализ повторных измерений (RM-ANOVA) и апостериорный тест Фишера для оценки различий в массе тела между экспериментальными методами лечения. Такую же методику применяли для второй опытной группы птиц, на которых оценивали состав тела. Мы оценили влияние наличия воды на BMR, TEWL, массы органов, биомаркеры окислительного стресса и концентрацию жидкости, используя общую линейную модель (GLM) с конечной массой тела в качестве ковариации и состоянием гидратации в качестве фиксированных факторов.Первоначально мы включили обработку по взаимодействию массы тела, но это взаимодействие не было значительным и было удалено из линейной модели. Затем мы снова проверили влияние ковариации массы тела и лечения, и когда ковариата оказалась незначимой, мы удалили этот член из модели. Чтобы оценить степень, в которой активность метаболических ферментов связана с массой тела и скоростью метаболизма, мы провели множественную линейную регрессию, используя общий BMR в качестве переменной ответа и активность ферментов и массу тела в качестве предикторов.Наконец, мы рассчитали и нанесли на график остатки линейной регрессии между массой тела и активностью ферментов. Все статистические анализы проводились с использованием статистического пакета STATISTICA ® для Windows. Для простоты здесь мы представляем результаты анализа с использованием ковариаты только тогда, когда GLM выявила значительное влияние массы тела. Подробные результаты влияния массы тела и взаимодействия между массой тела и лечением представлены в дополнительной таблице 1.
Затем мы снова проверили влияние ковариации массы тела и лечения, и когда ковариата оказалась незначимой, мы удалили этот член из модели. Чтобы оценить степень, в которой активность метаболических ферментов связана с массой тела и скоростью метаболизма, мы провели множественную линейную регрессию, используя общий BMR в качестве переменной ответа и активность ферментов и массу тела в качестве предикторов.Наконец, мы рассчитали и нанесли на график остатки линейной регрессии между массой тела и активностью ферментов. Все статистические анализы проводились с использованием статистического пакета STATISTICA ® для Windows. Для простоты здесь мы представляем результаты анализа с использованием ковариаты только тогда, когда GLM выявила значительное влияние массы тела. Подробные результаты влияния массы тела и взаимодействия между массой тела и лечением представлены в дополнительной таблице 1.
Результаты
Масса тела и органов
Мы обнаружили значительное влияние экспериментального времени на массу тела [RM ANOVA, F (3,48) = 47. 48; p <0,01], с более крутым снижением массы тела с течением времени в группе WR (рис. 1), так что к 7-му дню два лечения значительно отличались друг от друга [ F (1,16) = 25,82 ; p < 0,01, рис. 1]. Мы также обнаружили взаимосвязь времени и лечения с массой тела [ F (3,48) = 12,05; p < 0,01], при этом группа WR была наиболее поражена и потеряла в среднем до 17,7% исходной массы, тогда как группа CT уменьшилась только на 6.4% (рис. 1). Все массы внутренних органов коррелировали с конечной массой тела (см. Дополнительную таблицу 1B), и ANCOVAs показали, что при учете массы тела массы сердца [ F (1,15) = 5,67; p = 0,030] и почек [ F (1,15) = 9,621; p = 0,008] находились под влиянием ограничения воды, при этом группа WR имела более крупные органы, чем группа CT (таблица 1 и дополнительная таблица 1B).
48; p <0,01], с более крутым снижением массы тела с течением времени в группе WR (рис. 1), так что к 7-му дню два лечения значительно отличались друг от друга [ F (1,16) = 25,82 ; p < 0,01, рис. 1]. Мы также обнаружили взаимосвязь времени и лечения с массой тела [ F (3,48) = 12,05; p < 0,01], при этом группа WR была наиболее поражена и потеряла в среднем до 17,7% исходной массы, тогда как группа CT уменьшилась только на 6.4% (рис. 1). Все массы внутренних органов коррелировали с конечной массой тела (см. Дополнительную таблицу 1B), и ANCOVAs показали, что при учете массы тела массы сердца [ F (1,15) = 5,67; p = 0,030] и почек [ F (1,15) = 9,621; p = 0,008] находились под влиянием ограничения воды, при этом группа WR имела более крупные органы, чем группа CT (таблица 1 и дополнительная таблица 1B).
Рис. 1. Средняя масса тела Z. capensis , акклиматизированных к контрольным (CT, n = 9) и ограничению воды (WR, n = 9) условиям; планки погрешностей представляют собой стандартное отклонение. Буквы обозначают значительные различия между каждым лечением в определенные моменты времени с повторными измерениями ANOVA и апостериорным тестом Тьюки HSD.
capensis , акклиматизированных к контрольным (CT, n = 9) и ограничению воды (WR, n = 9) условиям; планки погрешностей представляют собой стандартное отклонение. Буквы обозначают значительные различия между каждым лечением в определенные моменты времени с повторными измерениями ANOVA и апостериорным тестом Тьюки HSD.
Таблица 1. Массы органов (среднее значение и LSM ± стандартная ошибка) Z. capensis , акклиматизированных к гидратации (контроль) и условиям ограничения воды (CT = контроль; n = 9 и WR = ограничение воды; n = 9).
Количественные магнитно-резонансные измерения
В конце второго эксперимента по акклиматизации птицы в группе WR имели ∼12% меньшую массу тела [ F (1,9) = 10,79; p < 0,001], снижение мышечной массы на 10% [ F (1,9) = 11,72; p = 0,008], и на 11% ниже общее содержание воды [ F (1,9) = 13,97; p = 0,005] по сравнению с группой КТ (табл. 2). Достоверных различий по жировой массе не обнаружено [ F (1,9) = 0.07; p = 0,79] и содержание свободной воды между обработками [ F (1,9) = 2,04; р = 0,18]. Рассчитанные значения индекса гидратации были на 2% выше в группе CT по сравнению с группой WR [ F (1,9) = 18,59; р = 0,002]. Наконец, мы обнаружили сильную и положительную связь между общим количеством воды в организме и безжировой массой, когда все люди были объединены ( r = 0,98; p < 0,001).
2). Достоверных различий по жировой массе не обнаружено [ F (1,9) = 0.07; p = 0,79] и содержание свободной воды между обработками [ F (1,9) = 2,04; р = 0,18]. Рассчитанные значения индекса гидратации были на 2% выше в группе CT по сравнению с группой WR [ F (1,9) = 18,59; р = 0,002]. Наконец, мы обнаружили сильную и положительную связь между общим количеством воды в организме и безжировой массой, когда все люди были объединены ( r = 0,98; p < 0,001).
Таблица 2. Среднее значение (±SE) состава тела у Z. capensis , акклиматизированных к условиям ограничения воды (WR) и контролю (CT) в течение 15 дней.
Физиологические реакции
Масса тела не влияла на концентрацию жидкостей организма (дополнительная таблица 1C). Осмоляльность плазмы крови существенно не различалась между экспериментальными обработками [ F (1,14) = 1,70; p = 0,21 Рис. 2], однако в конце 2-недельного периода акклиматизации мы наблюдали значительное увеличение ~57.0% [ F (1,14) = 123,2; p = 0,001] осмоляльности мочи при дегидратации по сравнению с гидратацией (рис. 2).
2], однако в конце 2-недельного периода акклиматизации мы наблюдали значительное увеличение ~57.0% [ F (1,14) = 123,2; p = 0,001] осмоляльности мочи при дегидратации по сравнению с гидратацией (рис. 2).
Рисунок 2. Осмоляльность мочи и плазмы (A) и ферментативная активность ЦОГ и ХС (B) в эритроцитах Z. capensis , акклиматизированных к контролю (CT) и ограничению воды (WR) ) условия. (C) показывает скорректированные средние значения активности CS после учета влияния массы тела.Звездочка обозначает значительную разницу между обеими группами после анализа GLM (см. Дополнительную таблицу 1A для получения подробной статистики). Данные представлены как среднее значение ± стандартная ошибка.
Воробьи в группе WR имели ∼12% более низкий TEWL всего организма по сравнению с птицами в группе CT [ F (1,16) = 5,42, p = 0,033], однако не было никакой разницы в BMR всего организма между экспериментальными обработками [ F (1,16) = 1,85, p = 0. 192; Таблица 2]. И BMR, и TEWL достоверно коррелировали с массой тела ( r 2 = 0,39; p = 0,005, r 2 = 0,282; p = 0,023 для BMR, 3LMR, 3 TEW, соответственно) хотя GLM показал, что только BMR значительно коррелировал с массой тела (дополнительная таблица 1A). Анализ GLM показал, что BMR с поправкой на массу (среднее наименьших квадратов, LSM) в группе WR был в среднем примерно на 12% выше, чем BMR в группе CT [ F (1,15) = 4.84; р = 0,044, табл. 3].
192; Таблица 2]. И BMR, и TEWL достоверно коррелировали с массой тела ( r 2 = 0,39; p = 0,005, r 2 = 0,282; p = 0,023 для BMR, 3LMR, 3 TEW, соответственно) хотя GLM показал, что только BMR значительно коррелировал с массой тела (дополнительная таблица 1A). Анализ GLM показал, что BMR с поправкой на массу (среднее наименьших квадратов, LSM) в группе WR был в среднем примерно на 12% выше, чем BMR в группе CT [ F (1,15) = 4.84; р = 0,044, табл. 3].
. масса тела у Z. capensis особей, акклиматизированных к условиям контроля (синий символ) и ограничения воды (красный символ). Линии указывают на существенные различия между гидратированными и дегидратированными группами.
Таблица 3. Средняя (±SE) и скорректированная по массе (наименьшие квадратичные средние, LSM) скорость основного обмена (BMR, mLO 2 ч –1 ) и общая потеря воды с испарением (TEWL, мг H 2 O h –1 ), предполагаемое метаболическое производство воды (МРМ, мг H 2 O ч –1 ), отношение MWP/TEWL и масса тела (Mb) Z. capensis , акклиматизированных к ограничению воды (WR) и контрольные (CT) условия.
capensis , акклиматизированных к ограничению воды (WR) и контрольные (CT) условия.
После поправки на массу тела активность ХС в эритроцитах была значительно выше при ХТ, чем при лечении ВР [ F (1,15) = 5.49, р = 0,030; Рисунок 2], а средняя активность метаболического фермента цитохром-с-оксидазы была значительно выше в группе WR, чем в группе CT [ F (1,16) = 26,08, p < 0,001, Фигуры 2 , 3 и дополнительная таблица 1A]. GLM-анализ объединенных данных показал, что на BMR всего животного влияла масса тела [ F (1,16) = 37,70, p < 0,0001], активность ЦОГ [ F (1,16) = 17.93, p = 0,001], но не по активности ХС [ F (1,16) = 0,10, p = 0,75]. Мы также наблюдали значительную и положительную корреляцию между остатками отношений между массой тела и BMR или активностью ЦОГ ( y = 279,0x + 0,00; r 2 = 0,560, p <0,001; рис. 4). .
4). .
Рисунок 4. Остаточные значения активности ТМВ и ЦОГ в зависимости от массы тела показывают положительную и достоверную корреляцию в Z.capensis акклиматизировался к условиям контроля (синий символ) и ограничению воды (красный символ).
Окислительный статус
Концентрации маркеров окислительного статуса значительно различались в разных тканях (рис. 5, 6), но масса тела не влияла на эти параметры, за исключением концентрации глутатиона в грудной мышце (см. Дополнительную таблицу 1D). Перекисное окисление липидов было выше в грудной мышце птиц WR [ F (1,16) = 6.98; p = 0,018, фиг.5А], а сердце [ F (1,16) = 2,44; p = 0,138], печень [ F (1,16) = 2,69; p = 0,127], плазма [ F (1,16) = 0,02; p = 0,870], а эритроциты [ F (1,16) = 1,73; p = 0,207] не показали достоверных различий между экспериментальными группами. Не наблюдалось существенных различий в концентрации перекиси водорода между обработками грудной мышцы [ F (1,16) = 0.61.; р = 0,446], сердце [ F (1,16) = 0,69; p = 0,419], печень [ F (1,16) = 1,03; p = 0,330], или плазма крови [ F (1,16) = 0,31; p = 0,586] (рис. 5В). TAC не отличался между обработками грудных мышц [ F (1,16) = 1,31; р = 0,259], сердце [ F (1,16) = 2,43; p = 0,139], или плазма крови [ F (1,16) = 0.24; р = 0,630; Рисунок 5С]. Грудные мышцы птиц WR имели более низкие концентрации глутатиона как для восстановленной формы GSH, так и для окисленной формы GSSH по сравнению с гидратированной группой (см. Дополнительную таблицу 1D), но это различие исчезло, когда была учтена масса тела [ F (1,15). ) = 0,17; р = 0,685 и F (1,16) = 0,34; p = 0,568 соответственно].
Не наблюдалось существенных различий в концентрации перекиси водорода между обработками грудной мышцы [ F (1,16) = 0.61.; р = 0,446], сердце [ F (1,16) = 0,69; p = 0,419], печень [ F (1,16) = 1,03; p = 0,330], или плазма крови [ F (1,16) = 0,31; p = 0,586] (рис. 5В). TAC не отличался между обработками грудных мышц [ F (1,16) = 1,31; р = 0,259], сердце [ F (1,16) = 2,43; p = 0,139], или плазма крови [ F (1,16) = 0.24; р = 0,630; Рисунок 5С]. Грудные мышцы птиц WR имели более низкие концентрации глутатиона как для восстановленной формы GSH, так и для окисленной формы GSSH по сравнению с гидратированной группой (см. Дополнительную таблицу 1D), но это различие исчезло, когда была учтена масса тела [ F (1,15). ) = 0,17; р = 0,685 и F (1,16) = 0,34; p = 0,568 соответственно]. Соотношение GSH/GSSG в грудной клетке между обработками не показало [ F (1,16) = 1.14; р = 0,302]. ГШ [ Ф (1,16) = 3,52; p = 0,080], GSSG [ F (1,16) = 0,93; p = 0,350] и GSH/GSSG [ F (1,16) = 2,43; p = 0,138] в печени не отличались между обработками (рис. 6).
Соотношение GSH/GSSG в грудной клетке между обработками не показало [ F (1,16) = 1.14; р = 0,302]. ГШ [ Ф (1,16) = 3,52; p = 0,080], GSSG [ F (1,16) = 0,93; p = 0,350] и GSH/GSSG [ F (1,16) = 2,43; p = 0,138] в печени не отличались между обработками (рис. 6).
Рисунок 5. Перекисное окисление липидов (A) , водные пероксиды (B) и общая антиоксидантная способность (C) , измеренная в грудной клетке, сердце, печени, плазме и эритроцитах (эритроцитах) в Z.capensis акклиматизировался к условиям контроля и ограничению воды. Данные представлены как среднее значение ± стандартная ошибка. Звездочки обозначают существенные различия между методами лечения.
Рисунок 6. Концентрации восстановленного и окисленного глутатиона (левая ось) и соотношение восстановленного/окисленного глутатиона (правая ось) в грудной клетке и печени Z. capensis , акклиматизированных к гидратированным и обезвоженным условиям. Данные представлены как среднее значение ± стандартная ошибка. Звездочки обозначают существенные различия между методами лечения.
capensis , акклиматизированных к гидратированным и обезвоженным условиям. Данные представлены как среднее значение ± стандартная ошибка. Звездочки обозначают существенные различия между методами лечения.
Обсуждение
Энергетическая и осморегулирующая коррекция
Некоторые виды птиц реагируют на нехватку (пресной) воды, увеличивая расход энергии (включая BMR), реакцию, которая обычно связана с повышенными осморегуляторными потребностями у птиц, населяющих засушливую среду или регулярно пьющих соленую воду (Gutiérrez et al., 2011; Сабат и др., 2017). Альтернативная точка зрения состоит в том, что такие реакции могут быть адаптивными, учитывая, что увеличение скорости метаболизма может привести к увеличению MWP, что снижает потребность в сохранении воды и/или зависимости от (заранее сформированной) питьевой/пищевой воды.В какой степени увеличение скорости метаболизма объясняется необходимостью производить большее количество метаболической воды, остается открытым вопросом. Недавние исследования, в которых количественно оценивались водный баланс у полевых и лабораторных животных, показали, что как внешние (например, температура акклиматизации), так и внутренние (масса тела, оцепенение) влияют на расход энергии, что, в свою очередь, может увеличить долю метаболической воды в общем организме. бассейн с водой (Cooper et al., 2019; Whiteman et al., 2019). Интересно, что морские птицы, которые регулярно пьют морскую воду, по-видимому, увеличивают скорость своего метаболизма на большее значение, чем это требуется для соответствия термодинамическим оценкам осмотической работы.Такое избыточное увеличение скорости метаболизма может быть адаптивным механизмом увеличения MWP для поддержания водного баланса (Peña-Villalobos et al., 2013; Sabat et al., 2017).
Недавние исследования, в которых количественно оценивались водный баланс у полевых и лабораторных животных, показали, что как внешние (например, температура акклиматизации), так и внутренние (масса тела, оцепенение) влияют на расход энергии, что, в свою очередь, может увеличить долю метаболической воды в общем организме. бассейн с водой (Cooper et al., 2019; Whiteman et al., 2019). Интересно, что морские птицы, которые регулярно пьют морскую воду, по-видимому, увеличивают скорость своего метаболизма на большее значение, чем это требуется для соответствия термодинамическим оценкам осмотической работы.Такое избыточное увеличение скорости метаболизма может быть адаптивным механизмом увеличения MWP для поддержания водного баланса (Peña-Villalobos et al., 2013; Sabat et al., 2017).
Обезвоживание и связанное с ним снижение массы тела (рис. 1) коррелируют с более высокими скорректированными по массе BMR и MWP у воробьев (таблица 3). В сочетании со снижением TEWL по мере снижения массы тела (рис. 3B) более высокий MWP, вероятно, сводил к минимуму обезвоживание при лечении WR. Наблюдаемая потеря массы тела и увеличение BMR с поправкой на массу могут свидетельствовать о том, что Z.capensis может одновременно усиливать катаболизм белков и выработку метаболической воды посредством окисления (Gerson and Guglielmo, 2011). Это может свести к минимуму уровень обезвоживания тканей, поскольку большинство тканей имели одинаковые массы независимо от гидратации (таблица 1). Хотя эти корректировки могут быть адаптивными и привести к большей устойчивости к ограниченной доступности воды, эта стратегия является дорогостоящей с точки зрения потери массы. У птиц катаболизм белковой ткани обеспечивает более высокое чистое количество предварительно образованной воды на грамм по сравнению с катаболизмом липидов (Gerson and Guglielmo, 2011), поэтому способность предпочтительно использовать белок вместо запасов липидов может помочь поддерживать водный баланс в организме. за счет потери мышц и, следовательно, мышечной массы тела.
3B) более высокий MWP, вероятно, сводил к минимуму обезвоживание при лечении WR. Наблюдаемая потеря массы тела и увеличение BMR с поправкой на массу могут свидетельствовать о том, что Z.capensis может одновременно усиливать катаболизм белков и выработку метаболической воды посредством окисления (Gerson and Guglielmo, 2011). Это может свести к минимуму уровень обезвоживания тканей, поскольку большинство тканей имели одинаковые массы независимо от гидратации (таблица 1). Хотя эти корректировки могут быть адаптивными и привести к большей устойчивости к ограниченной доступности воды, эта стратегия является дорогостоящей с точки зрения потери массы. У птиц катаболизм белковой ткани обеспечивает более высокое чистое количество предварительно образованной воды на грамм по сравнению с катаболизмом липидов (Gerson and Guglielmo, 2011), поэтому способность предпочтительно использовать белок вместо запасов липидов может помочь поддерживать водный баланс в организме. за счет потери мышц и, следовательно, мышечной массы тела. Скорость метаболизма всего животного была одинаковой между экспериментальными обработками (таблица 3), несмотря на значительное снижение массы тела, наблюдаемое в группе WR. Возможно, что если наблюдаемое снижение массы тела было в основном связано с потерей воды, а не с потерей мышечной массы, то мы переоцениваем BMR с поправкой на массу, поскольку вода в организме не участвует напрямую в метаболизме. К сожалению, мы не смогли измерить изменения в составе тела до и после первоначального эксперимента, но второе испытание с акклиматизацией позволило нам оценить, была ли потеря массы тела при лечении WR обусловлена изменениями мышечной массы, содержания жира или воды.Минимальная (2%) разница в индексе гидратации, которую мы наблюдали между обработками (около 0,85, таблица 2), в сочетании с наблюдением, что изменения в общем содержании воды были связаны с уменьшением нежировой массы, предполагает, что наблюдаемое снижение общего содержания воды, вероятно, было следствие потери мышечной массы (Guglielmo et al.
Скорость метаболизма всего животного была одинаковой между экспериментальными обработками (таблица 3), несмотря на значительное снижение массы тела, наблюдаемое в группе WR. Возможно, что если наблюдаемое снижение массы тела было в основном связано с потерей воды, а не с потерей мышечной массы, то мы переоцениваем BMR с поправкой на массу, поскольку вода в организме не участвует напрямую в метаболизме. К сожалению, мы не смогли измерить изменения в составе тела до и после первоначального эксперимента, но второе испытание с акклиматизацией позволило нам оценить, была ли потеря массы тела при лечении WR обусловлена изменениями мышечной массы, содержания жира или воды.Минимальная (2%) разница в индексе гидратации, которую мы наблюдали между обработками (около 0,85, таблица 2), в сочетании с наблюдением, что изменения в общем содержании воды были связаны с уменьшением нежировой массы, предполагает, что наблюдаемое снижение общего содержания воды, вероятно, было следствие потери мышечной массы (Guglielmo et al. , 2011). Действительно, наши результаты показывают, что птицы WR увеличили катаболизм белка, потому что их безжировая масса уменьшилась примерно на 10%, что согласуется с общим снижением массы тела примерно на 12%.Таким образом, в то время как более низкое содержание воды в тканях птиц может частично объяснить более высокий скорректированный по массе BMR у птиц WR, большая часть изменений BMR, вероятно, обусловлена увеличением удельной метаболической активности внутренних органов (сердца и почек) и мышечной ткани. . Поскольку белок катаболизируется по-разному между двумя экспериментальными группами, но BMR у целых животных не различался, наблюдаемые изменения в масс-специфическом BMR позволяют предположить, что метаболическая активность конкретных тканей достаточно пластична (см. ниже).Тем не менее, мы предупреждаем, что BMR и состояние тела измерялись у разных людей, поэтому эта гипотеза является предварительной и требует дальнейшего изучения.
, 2011). Действительно, наши результаты показывают, что птицы WR увеличили катаболизм белка, потому что их безжировая масса уменьшилась примерно на 10%, что согласуется с общим снижением массы тела примерно на 12%.Таким образом, в то время как более низкое содержание воды в тканях птиц может частично объяснить более высокий скорректированный по массе BMR у птиц WR, большая часть изменений BMR, вероятно, обусловлена увеличением удельной метаболической активности внутренних органов (сердца и почек) и мышечной ткани. . Поскольку белок катаболизируется по-разному между двумя экспериментальными группами, но BMR у целых животных не различался, наблюдаемые изменения в масс-специфическом BMR позволяют предположить, что метаболическая активность конкретных тканей достаточно пластична (см. ниже).Тем не менее, мы предупреждаем, что BMR и состояние тела измерялись у разных людей, поэтому эта гипотеза является предварительной и требует дальнейшего изучения.
Изменение скорости метаболизма у птиц обычно связано с изменениями удельных энергозатрат внутренних органов, на которые, в свою очередь, влияет как масса органов, так и массоспецифический метаболизм тканей, контролируемый плотностью митохондрий и/или активностью ферментов (Vezina and Williams, 2005). ; Сабат и др., 2019). Результаты для Z.capensis подтверждают идею о том, что обезвоживание уменьшает размеры тела, что приводит к увеличению относительного размера почек и сердца (табл. 1). Похоже, что увеличение осморегуляторной потребности, связанное с необходимостью концентрировать мочу у некоторых видов воробьиных, потребляющих богатые белком и соленые ресурсы, вызывает одинаковую реакцию в морфологии почек и сердца (например, Sabat et al., 2004a,b; Peña- Виллалобос и др., 2013). Это интересно, так как сообщалось, что эти внутренние органы имеют непропорционально высокую скорость метаболизма, специфичную для массы, и что вариации независимого от массы метаболизма отражают относительный размер этого метаболически активного механизма (Daan et al., 1990). К сожалению, мы измерили только удельную скорость метаболизма крови, которая была значительно выше в группе WR, чем в группе CT. Несмотря на то, что объем крови составляет ∼8–10% от общего объема у птиц (Sturkie and Griminger, 1986), неизвестно, какой пропорциональный вклад клетки крови вносят в общую скорость метаболизма.
; Сабат и др., 2019). Результаты для Z.capensis подтверждают идею о том, что обезвоживание уменьшает размеры тела, что приводит к увеличению относительного размера почек и сердца (табл. 1). Похоже, что увеличение осморегуляторной потребности, связанное с необходимостью концентрировать мочу у некоторых видов воробьиных, потребляющих богатые белком и соленые ресурсы, вызывает одинаковую реакцию в морфологии почек и сердца (например, Sabat et al., 2004a,b; Peña- Виллалобос и др., 2013). Это интересно, так как сообщалось, что эти внутренние органы имеют непропорционально высокую скорость метаболизма, специфичную для массы, и что вариации независимого от массы метаболизма отражают относительный размер этого метаболически активного механизма (Daan et al., 1990). К сожалению, мы измерили только удельную скорость метаболизма крови, которая была значительно выше в группе WR, чем в группе CT. Несмотря на то, что объем крови составляет ∼8–10% от общего объема у птиц (Sturkie and Griminger, 1986), неизвестно, какой пропорциональный вклад клетки крови вносят в общую скорость метаболизма. Норд и др. (2021) сообщили, что каменные синицы ( Periparus ater ) и большие синицы ( Parus major ) увеличивают частоту митохондриального дыхания и активность CS эритроцитов зимой, что может быть ответом на изменения метаболических потребностей между сезонами.В следующих разделах мы предлагаем альтернативные объяснения наблюдаемого увеличения BMR с поправкой на массу и активности ЦОГ в эритроцитах и почему это может быть связано с затратами осмотического стресса.
Норд и др. (2021) сообщили, что каменные синицы ( Periparus ater ) и большие синицы ( Parus major ) увеличивают частоту митохондриального дыхания и активность CS эритроцитов зимой, что может быть ответом на изменения метаболических потребностей между сезонами.В следующих разделах мы предлагаем альтернативные объяснения наблюдаемого увеличения BMR с поправкой на массу и активности ЦОГ в эритроцитах и почему это может быть связано с затратами осмотического стресса.
Изменения метаболической активности эритроцитов
Возможно, самым примечательным результатом нашего исследования стало то, что у птиц наблюдалось повышение активности ЦОГ в эритроцитах при хроническом обезвоживании по сравнению с птицами, имевшими свободный доступ к воде. Кроме того, изменение активности ЦОГ эритроцитов было связано с изменением уровня расхода энергии.Одной из уникальных характеристик птиц, которую экологи и эволюционные физиологи часто упускают из виду, является наличие ядерных эритроцитов с функциональными митохондриями. Эта ключевая особенность недавно привлекла внимание физиологов, и существование внутри- и межвидовой изменчивости ее функции наблюдалось у нескольких видов (Stier et al., 2013, 2015, 2017; Nord et al., 2021). Несколько десятилетий назад некоторые предполагали, что продукция АТФ в эритроцитах участвует в поддержании баланса ионов (Na и K) между эритроцитами и плазмой (Hunter and Hunter, 1957).Другие предположили, что наличие ядерных эритроцитов с функциональными митохондриями может быть связано с механизмами регуляции объема клетки с использованием белков переноса ионов (например, Na + /H + , Na + -K + -2Cl – котранспорт и каналы Na + ), которые опосредуются АТФ-зависимыми мембранными белками (например, NHEs, KCC и NKCC; Hoffmann et al., 2009). Однако наши результаты для Z. capensis не обязательно подтверждают гипотезу о том, что увеличение митохондриальной окислительной активности в эритроцитах обусловлено исключительно регуляцией клеточного объема.
Эта ключевая особенность недавно привлекла внимание физиологов, и существование внутри- и межвидовой изменчивости ее функции наблюдалось у нескольких видов (Stier et al., 2013, 2015, 2017; Nord et al., 2021). Несколько десятилетий назад некоторые предполагали, что продукция АТФ в эритроцитах участвует в поддержании баланса ионов (Na и K) между эритроцитами и плазмой (Hunter and Hunter, 1957).Другие предположили, что наличие ядерных эритроцитов с функциональными митохондриями может быть связано с механизмами регуляции объема клетки с использованием белков переноса ионов (например, Na + /H + , Na + -K + -2Cl – котранспорт и каналы Na + ), которые опосредуются АТФ-зависимыми мембранными белками (например, NHEs, KCC и NKCC; Hoffmann et al., 2009). Однако наши результаты для Z. capensis не обязательно подтверждают гипотезу о том, что увеличение митохондриальной окислительной активности в эритроцитах обусловлено исключительно регуляцией клеточного объема. Осмоляльность плазмы была одинаковой у птиц WR и CT, и, в более широком смысле, осмотическое давление, испытываемое эритроцитами, вероятно, было одинаковым между обработками, что также позволяет предположить, что Zonotrichia была способна поддерживать водный гомеостаз в нашем эксперименте.
Осмоляльность плазмы была одинаковой у птиц WR и CT, и, в более широком смысле, осмотическое давление, испытываемое эритроцитами, вероятно, было одинаковым между обработками, что также позволяет предположить, что Zonotrichia была способна поддерживать водный гомеостаз в нашем эксперименте.
Хотя отсутствие опубликованных данных не позволяет дать четкое объяснение наблюдаемой вариации активности ЦОГ и ХС в эритроцитах, интересно, что после исключения влияния массы тела только активность ЦОГ положительно коррелирует с BMR.Этот вывод согласуется с теми, о которых недавно сообщили Gutiérrez et al. (2019) для гудзоновского веретенника ( Limosa haemastica ), которые предположили, что скорость расхода энергии у веретенника может зависеть от метаболических потребностей органов и тканей, включая кровь. Хотя наши результаты показывают несколько противоречивые закономерности экспрессии ЦОГ и ХС в эритроцитах, сопоставимые результаты также наблюдались у гудзоновских веретенников, собранных зимой (Gutiérrez et al. , 2019). Как указали эти авторы, могут существовать и другие физиологические процессы, такие как рост и восстановление тканей, которые по-разному модулируют экспрессию ХС и ЦОГ в эритроцитах птиц (Velando et al., 2019).
, 2019). Как указали эти авторы, могут существовать и другие физиологические процессы, такие как рост и восстановление тканей, которые по-разному модулируют экспрессию ХС и ЦОГ в эритроцитах птиц (Velando et al., 2019).
Для Z. capensis по крайней мере две неисключительные альтернативы могут объяснить взаимосвязь, обнаруженную между скорректированным по массе BMR и активностью ЦОГ эритроцитов. Во-первых, увеличение катаболической способности эритроцитов может быть реакцией на вазоконстрикцию, вызванную ограничением воды. Хорошо известно, что обезвоживание вызывает сужение сосудов кровеносных капилляров, что снижает доставку кислорода к тканям (Kreimeier, 2000). Кроме того, эритроциты могут определять напряжение кислорода посредством микроциркуляции, регулируя сосудистый тонус капилляров в определенных тканях (Jensen, 2009).В случае гипоксии, вызванной вазоконстрикцией, вызванной обезвоживанием, эритроциты могут увеличивать высвобождение сосудорасширяющих соединений (например, NO, АТФ), локально способствуя увеличению перфузии кровью тканей, нуждающихся в кислороде (Ellsworth et al. , 1995) и, таким образом, повышает его окислительную активность. Во-вторых, вполне вероятно, что изменения метаболической активности в крови могут быть вызваны усилением тканевого метаболизма. Пищевые датчики или белки, которые реагируют на колебания питательных веществ, такие как никотинамидфосфорибозилтрансфераза, сиртуины (молчаливый информационный регулятор) и мишень рапамицинового комплекса, вероятно, зависят от энергетического статуса организма.Столкнувшись с высокой потребностью в кислороде, животные реагируют увеличением потребления питательных веществ тканями с более высокой удельной массой метаболизма, такими как почки (Gaur et al., 2017; Lee and Yang, 2017), что, вероятно, влияет на другие ткани. Таким образом, наблюдаемая взаимосвязь между BMR и ЦОГ в эритроцитах может отражать синергический ответ на поддержание сосудистого и дыхательного гомеостаза либо потому, что клетки крови вносят вклад в регулирование кровотока сосудистого тонуса, либо потому, что на них влияют те же регуляторные пути, которые отражают потребности других энергоемких тканей (например,г.
, 1995) и, таким образом, повышает его окислительную активность. Во-вторых, вполне вероятно, что изменения метаболической активности в крови могут быть вызваны усилением тканевого метаболизма. Пищевые датчики или белки, которые реагируют на колебания питательных веществ, такие как никотинамидфосфорибозилтрансфераза, сиртуины (молчаливый информационный регулятор) и мишень рапамицинового комплекса, вероятно, зависят от энергетического статуса организма.Столкнувшись с высокой потребностью в кислороде, животные реагируют увеличением потребления питательных веществ тканями с более высокой удельной массой метаболизма, такими как почки (Gaur et al., 2017; Lee and Yang, 2017), что, вероятно, влияет на другие ткани. Таким образом, наблюдаемая взаимосвязь между BMR и ЦОГ в эритроцитах может отражать синергический ответ на поддержание сосудистого и дыхательного гомеостаза либо потому, что клетки крови вносят вклад в регулирование кровотока сосудистого тонуса, либо потому, что на них влияют те же регуляторные пути, которые отражают потребности других энергоемких тканей (например,г. , печень). Малкок и др. (2021) сообщили, что потребление кислорода на клеточном уровне в эритроцитах больших синиц ( Parus major ) напрямую связано с потреблением кислорода на уровне организма. Эта закономерность наблюдалась только тогда, когда птицы не проявляли признаков стресса, как следует из концентрации кортикостерона в плазме, что указывает на то, что работа по изучению влияния стресса на скорость метаболизма как на митохондриальном, так и на организменном уровне требует дальнейшего изучения.
, печень). Малкок и др. (2021) сообщили, что потребление кислорода на клеточном уровне в эритроцитах больших синиц ( Parus major ) напрямую связано с потреблением кислорода на уровне организма. Эта закономерность наблюдалась только тогда, когда птицы не проявляли признаков стресса, как следует из концентрации кортикостерона в плазме, что указывает на то, что работа по изучению влияния стресса на скорость метаболизма как на митохондриальном, так и на организменном уровне требует дальнейшего изучения.
Окислительный статус
Воробьи в группе WR имели более высокие уровни перекисного окисления липидов, измеренные по концентрации МДА в грудных мышцах, что является признаком более высокого окислительного повреждения по сравнению с птицами, имевшими свободный доступ к воде.Эти данные подтверждаются нашей предыдущей работой, показывающей увеличение концентрации МДА на 400% в Z. capensis , получавших соленую воду (Sabat et al., 2017). Наблюдаемые различия в окислительном повреждении грудной мышцы между обработками в этом эксперименте предполагают наличие признаков окислительного стресса у Z. capensis , подвергшихся ограничению воды, несмотря на то, что ни прооксиданты (пероксиды), ни антиоксидантная способность (ТАС) не различаются между обработками. Повреждение мембран в грудных мышцах обезвоженных птиц может быть частично объяснено наблюдаемым увеличением удельного BMR, что может способствовать выработке свободных радикалов или АФК.В исследованиях сообщалось о противоречивых результатах относительно функциональной связи между скоростью метаболизма и продукцией АФК в контролируемых условиях как при экзотермах, так и при эндотермах (Hermes-Lima and Storey, 1998; Brzȩ et al., 2014; Salin et al., 2015; Zhang and Wong, 2021). К сожалению, в нескольких исследованиях оценивались последствия обезвоживания и стресса (включая окислительное повреждение) у птиц, хотя в недавней публикации сообщалось о связи между обезвоживанием и повышением концентрации кортикостерона у домашних воробьев, живущих на свободном выгуле ( Passer domesticus ; Brischoux et al., 2020). Хотя известно, что кортикостерон и другие глюкокортикоиды нарушают клеточный окислительный гомеостаз (Costantini et al.
capensis , подвергшихся ограничению воды, несмотря на то, что ни прооксиданты (пероксиды), ни антиоксидантная способность (ТАС) не различаются между обработками. Повреждение мембран в грудных мышцах обезвоженных птиц может быть частично объяснено наблюдаемым увеличением удельного BMR, что может способствовать выработке свободных радикалов или АФК.В исследованиях сообщалось о противоречивых результатах относительно функциональной связи между скоростью метаболизма и продукцией АФК в контролируемых условиях как при экзотермах, так и при эндотермах (Hermes-Lima and Storey, 1998; Brzȩ et al., 2014; Salin et al., 2015; Zhang and Wong, 2021). К сожалению, в нескольких исследованиях оценивались последствия обезвоживания и стресса (включая окислительное повреждение) у птиц, хотя в недавней публикации сообщалось о связи между обезвоживанием и повышением концентрации кортикостерона у домашних воробьев, живущих на свободном выгуле ( Passer domesticus ; Brischoux et al., 2020). Хотя известно, что кортикостерон и другие глюкокортикоиды нарушают клеточный окислительный гомеостаз (Costantini et al. , 2011; Queisser et al., 2011), неизвестно, вызывает ли обезвоживание окислительное повреждение у птиц. Необходимы дополнительные экспериментальные данные для точной оценки того, как обезвоживание влияет на окислительный статус у птиц и какие внутренние (например, биохимические) и/или внешние (например, экологические) механизмы лежат в основе их реакции.
, 2011; Queisser et al., 2011), неизвестно, вызывает ли обезвоживание окислительное повреждение у птиц. Необходимы дополнительные экспериментальные данные для точной оценки того, как обезвоживание влияет на окислительный статус у птиц и какие внутренние (например, биохимические) и/или внешние (например, экологические) механизмы лежат в основе их реакции.
Заключение
Мы нашли пленника Z.capensis реагируют на ограничение воды, теряя массу и увеличивая свой метаболизм, зависящий от массы тела, модель, которая предполагает, что одновременное увеличение катаболизма белка и производство метаболической воды посредством окисления может помочь поддерживать водный баланс. Примечательным результатом является то, что у птиц наблюдалось увеличение активности ЦОГ в эритроцитах в ответ на хроническое ограничение воды, и что изменение активности ЦОГ положительно коррелировало с BMR с поправкой на массу. Наконец, мы обнаружили умеренное влияние ограничения воды на повреждение мембран скелетных мышц, что подтверждает, что состояние гидратации и изменения осмотических условий могут вызывать окислительное повреждение. Необходимо дальнейшее изучение того, как птицы используют физиологические механизмы для поддержания водного баланса, чтобы лучше понять, как они будут реагировать на быстро меняющиеся условия окружающей среды, которые, вероятно, вызовут повышение температуры окружающей среды, но снижение доступности воды в экосистемах засушливых земель во всем мире.
Необходимо дальнейшее изучение того, как птицы используют физиологические механизмы для поддержания водного баланса, чтобы лучше понять, как они будут реагировать на быстро меняющиеся условия окружающей среды, которые, вероятно, вызовут повышение температуры окружающей среды, но снижение доступности воды в экосистемах засушливых земель во всем мире.
Заявление о доступности данных
Необработанные данные, подтверждающие выводы этой статьи, будут предоставлены авторами без неоправданных оговорок.
Заявление об этике
Исследование на животных было рассмотрено и одобрено Институциональным комитетом по уходу за животными Чилийского университета.
Вклад авторов
Исследования PS, SN, FB и RN. LN, IP-V, CC-R и JS-H провели исследования. LN и PS проанализировали данные. PS, SN, FB, RN, JS-H, LN и IP-V написали рукопись. Все авторы внесли свой вклад в статью и одобрили представленную версию.
Финансирование
Эта работа финансировалась ANID/FONDECYT Regular N° 1200386, ANID PIA/BASAL FB0002 и грантом FONDEQUIP N° EQM180055.
Конфликт интересов
Авторы заявляют, что исследование проводилось при отсутствии каких-либо коммерческих или финансовых отношений, которые могли бы быть истолкованы как потенциальный конфликт интересов.
Примечание издателя
Все претензии, изложенные в этой статье, принадлежат исключительно авторам и не обязательно представляют претензии их дочерних организаций или издателя, редакторов и рецензентов. Любой продукт, который может быть оценен в этой статье, или претензии, которые могут быть сделаны его производителем, не гарантируются и не поддерживаются издателем.
Благодарности
Мы благодарим Felipe Alvarez, Carlos Mejias, Andrés Sazo и Natalia Ramirez за неоценимую помощь в лабораторных и полевых работах.
Дополнительный материал
Дополнительный материал к этой статье можно найти в Интернете по адресу: https://www.frontiersin.org/articles/10.3389/fevo.2021.767280/full#supplementary-material
.Каталожные номера
Олбрайт, Т. П., Мутибва, Д., Герсон, А. Р., Смит, Э. К., Талбот, В. А., О’Нил, Дж.Дж. и др. (2017). Картирование потери воды при испарении пустынными воробьиными показывает растущую угрозу смертельного обезвоживания. Проц. Натл. акад. науч. США 114, 2283–2288. doi: 10.1073/pnas.1613625114
П., Мутибва, Д., Герсон, А. Р., Смит, Э. К., Талбот, В. А., О’Нил, Дж.Дж. и др. (2017). Картирование потери воды при испарении пустынными воробьиными показывает растущую угрозу смертельного обезвоживания. Проц. Натл. акад. науч. США 114, 2283–2288. doi: 10.1073/pnas.1613625114
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Брэдфорд, Массачусетс (1976). Быстрый и чувствительный метод количественного определения белка в микрограммах, использующий принцип связывания белка с красителем. Анал. Биохим. 72, 248–254. дои: 10.1016/0003-2697(76)-3
Полнотекстовая перекрестная ссылка | Академия Google
Бришу, Ф., Божеар, Э., Моринг, Б., Паренто, К., и Анжелье, Ф. (2020). Кратковременное обезвоживание влияет на исходный, но не на вызванный стрессом уровень кортикостерона у домашнего воробья ( Passer domesticus ). Дж. Экспл. биол. 223:jeb216424. doi: 10.1242/jeb.216424
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Бжцк, П. , Ксяжек, А., Олдаковски, Л. и Конаржевски, М.(2014). Высокая скорость основного обмена не вызывает окислительного стресса во время размножения у лабораторных мышей. Дж. Экспл. биол. 217, 1504–1509. doi: 10.1242/jeb.100073
, Ксяжек, А., Олдаковски, Л. и Конаржевски, М.(2014). Высокая скорость основного обмена не вызывает окислительного стресса во время размножения у лабораторных мышей. Дж. Экспл. биол. 217, 1504–1509. doi: 10.1242/jeb.100073
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Кейд, Т.Дж., Тобин, К.А., и Голд, А. (1965). Водное хозяйство и метаболизм двух эстридиновых вьюрков. Физиол. Зоол. 38, 9–33. doi: 10.1086/physzool.38.1.30152342
Полнотекстовая перекрестная ссылка | Академия Google
Коэн, А.А., Хау М. и Викельски М. (2008). Стресс, обмен веществ и антиоксиданты у двух видов диких воробьиных птиц. Физиол. Биохим. Зоол . 81, 463–472. дои: 10.1086/589548
Полнотекстовая перекрестная ссылка | Академия Google
Купер, К.Э., Уизерс, П.К., Херли, Л.Л., и Гриффит, С.К. (2019). Скорость метаболизма в полевых условиях, круговорот воды, пищевое и питьевое поведение мелких птичьих зерноядных пустынь во время летней жары. Фронт. Физиол. 10:1405. дои: 10.3389/ффиз.2019.01405
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Костантини, Д. (2014). Окислительный стресс и гормезис в эволюционной экологии и физиологии: сочетание механистического и эволюционного подходов. Берлин: Springer-Verlag. дои: 10.1007/978-3-642-54663-1
Полнотекстовая перекрестная ссылка | Академия Google
Костантини, Д., Мараско, В., и Меллер, А. П. (2011). Метаанализ глюкокортикоидов как модуляторов окислительного стресса у позвоночных. Дж. Комп. Физиол. Б. 181, 447–456. doi: 10.1007/s00360-011-0566-2
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Костантини Д., Роу М., Батлер М. В. и Макгроу К. Дж. (2010). От молекул к живым системам: исторические и современные проблемы окислительного стресса и экологии антиоксидантов. Функц. Экол. 24, 950–959. doi: 10.1111/j.1365-2435.2010.01746.x
Полнотекстовая перекрестная ссылка | Академия Google
Даан С. , Масман Д.и Гроэневолд, А. (1990). Основные скорости метаболизма птиц: их связь с составом тела и расходом энергии в природе. утра. Дж. Физ. Регул. интегр. Комп. физ. 259, Р333–Р340. doi: 10.1152/ajpregu.1990.259.2.R333
, Масман Д.и Гроэневолд, А. (1990). Основные скорости метаболизма птиц: их связь с составом тела и расходом энергии в природе. утра. Дж. Физ. Регул. интегр. Комп. физ. 259, Р333–Р340. doi: 10.1152/ajpregu.1990.259.2.R333
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Истик, Д.Л., Эдвардс, А.М., Гриффитс, С.Р., Спенсер, С.Дж., и Роберт, К.А. (2020). Валидация количественного магнитного резонанса как неинвазивного измерения состава тела у австралийской летучей мыши. австр. Млекопитающее. 43, 196–202. дои: 10.1071/AM19060
Полнотекстовая перекрестная ссылка | Академия Google
Эллсуорт, М.Л., Форрестер, Т., Эллис, К.Г., и Дитрих, Х.Х. (1995). Эритроцит как регулятор сосудистого тонуса. утра. Дж. Физиол. Цирк Сердца. Физиол. 269, h3155–h3161. doi: 10.1152/ajpheart.1995.269.6.h3155
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Гарднер, Дж. Л., Питерс, А., Кирни, М. Р., Джозеф, Л., и Хейнсон, Р.(2011). Уменьшение размеров тела: третий универсальный ответ на потепление? Тренды Экол. Эвол. 26, 285–291. doi: 10.1016/j.tree.2011.03.005
Р., Джозеф, Л., и Хейнсон, Р.(2011). Уменьшение размеров тела: третий универсальный ответ на потепление? Тренды Экол. Эвол. 26, 285–291. doi: 10.1016/j.tree.2011.03.005
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Gaur, U., Tu, J., Li, D., Gao, Y., Lian, T., Sun, B., et al. (2017). Молекулярно-эволюционные паттерны сигнального пути старения НАД+/сиртуин для разных таксонов. PLoS One 12:e0182306. doi: 10.1371/journal.pone.0182306
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Герсон, А.Р. и Гульельмо, К.Г. (2011). Домовые воробьи ( Passer domesticus ) усиливают катаболизм белка в ответ на ограничение воды. утра. Дж. Физиол. Регул. интегр. Комп. Физиол. 300, R925–R930. doi: 10.1152/ajpregu.00701.2010
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Гульельмо, К.Г., Макгуайр, Л.П., Герсон, А.Р., и Сиваген, К.Л. (2011). Простое, быстрое и неинвазивное измерение массы жира, мяса и общей массы воды у живых птиц с помощью количественного магнитного резонанса. Дж. Орнитол. 152:75. doi: 10.1007/s10336-011-0724-z
Дж. Орнитол. 152:75. doi: 10.1007/s10336-011-0724-z
Полнотекстовая перекрестная ссылка | Академия Google
Гутьеррес, Дж. С., Масеро, Дж. А., Абад-Гомес, Дж. М., Вильегас, А., и Санчес-Гусман, Дж. М. (2011). Понимание энергетических затрат на жизнь в соленой среде: влияние солености на скорость основного обмена, массу тела и ежедневное потребление энергии куликов, мигрирующих на большие расстояния. Дж. Экспл. биол. 214, 829–835. doi: 10.1242/jeb.048223
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Гутьеррес, Х.С., Сабат П., Кастанеда Л.Э., Контрерас К., Наваррете Л., Пенья-Вильялобос И. и др. (2019). Окислительный статус и метаболический профиль у долгоживущих птиц, готовящихся к экстремальной миграции. Науч. Респ. 9:17616. doi: 10.1038/s41598-019-54057-6
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Гермес-Лима, М., и Стори, К.Б. (1998). Роль антиоксидантной защиты в переносимости тяжелой дегидратации бесхвостыми животными. случай с леопардовой лягушкой Rana pipiens . Мол. Клетка. Биохим. 189, 79–89. дои: 10.1023/A:1006868208476
случай с леопардовой лягушкой Rana pipiens . Мол. Клетка. Биохим. 189, 79–89. дои: 10.1023/A:1006868208476
Полнотекстовая перекрестная ссылка | Академия Google
Икнаян, К. Дж., и Бейссингер, С. Р. (2018). Крах сообщества пустынных птиц за последнее столетие, вызванный изменением климата. ПНАС 115, 8597–8602. doi: 10.1073/pnas.1805123115
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Джейкобс, П.Дж., Остуизен, М.К., Митчелл, К., Блаунт, Дж.Д., и Беннетт, Н.К.(2020). Тепло и обезвоживание вызывали окислительное повреждение и антиоксидантную защиту после теплового стресса в инкубаторе и смоделированной волны тепла у пойманных в дикой природе четырехполосых полевых мышей Rhabdomys dilectus . PLoS One 15:e0242279. doi: 10.1371/journal.pone.0242279
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Дженсен, Ф. Б. (2009). Двойная роль эритроцитов в доставке кислорода тканям: переносчиков кислорода и регуляторов местного кровотока. Дж.Эксп. биол. 212, 3387–3393. doi: 10.1242/jeb.023697
Дж.Эксп. биол. 212, 3387–3393. doi: 10.1242/jeb.023697
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Келси, Н. А., и Байрлейн, Ф. (2019). Мигрирующее увеличение массы тела северных каменок ( Oenanthe oenanthe ) связано с накоплением жира, что подтверждается количественным магнитным резонансом. Дж. Орнитол. 160, 389–397. doi: 10.1007/s10336-018-1621-5
Полнотекстовая перекрестная ссылка | Академия Google
Халик, И., Хоф, К., Принцингер, Р., Бенинг-Газе, К., и Пфеннингер, М. (2014). Глобальные различия в термостойкости и уязвимость эндотермов к изменению климата. Проц. Р. Соц. Б: биол. науч. 281:20141097. doi: 10.1098/rspb.2014.1097
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Кинг, Дж. Р., и Фарнер, Д. С. (1961). «Энергетический обмен, терморегуляция и температура тела», в Биология и сравнительная физиология птиц , изд. А. Дж. Маршалл (Нью-Йорк, Нью-Йорк: Academic Press).
Академия Google
Крафт, Ф.Л.О., Дрисколл, С.К., Бьюкенен, К.Л., и Крино, О.Л. (2019). Стресс, связанный с развитием, снижает состояние организма на разных этапах жизни птиц: сравнение количественных данных магнитного резонанса и индексов состояния. Ген. комп. Эндокринол. 272, 33–41. doi: 10.1016/j.ygcen.2018.11.008
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Ли, Х.Дж., и Ян, С.Дж. (2017). Связанная со старением корреляция между активностью сиртуина 1 в сыворотке крови и скоростью основного обмена у женщин, но не у мужчин. клин. Нутр. Рез. 6, 18–26. doi: 10.7762/cnr.2017.6.1.18
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Лиде, Д. Р. (2001). CRC Справочник по физике и химии. Бока-Ратон, Флорида: CRC Press.
Академия Google
Мальдонадо, К. Э., Кавьерес, Г., Велозу, К., Каналс, М., и Сабат, П. (2009). Физиологические реакции красношейных воробьев на температурную акклиматизацию и сезонную акклиматизацию. Дж. Комп. Физиол.В 179, 335–343. doi: 10.1007/s00360-008-0317-1
Дж. Комп. Физиол.В 179, 335–343. doi: 10.1007/s00360-008-0317-1
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
МакКечни, А.Э., и Вольф, Б.О. (2010). Изменение климата увеличивает вероятность катастрофической гибели птиц во время экстремальной жары. Биол. лат. 6, 253–256. doi: 10.1098/rsbl.2009.0702
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Маквортер Т.Дж., дель Рио К.М., Пиншоу Б. и Роксбург Л.(2004). Функция почек у палестинских нектарников: выведение избытка воды не ограничивает потребление энергии. Дж. Экспл. биол. 207, 3391–3398. doi: 10.1242/jeb.01169
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Мориц, К., Лим, Г.Б., и Винтур, Э. (1997). Регуляция развития эритропоэтина и эритропоэза. утра. Дж. Физиол. Регул. интегр. Комп. Физиол. 273, R1829–R1844. doi: 10.1152/ajpregu.1997.273.6.R1829
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Норд, А. , Меткалф, Н.Б., Пейдж, Дж.Л., Хакстейбл, А., Маккафферти, Д.Дж., и Доусон, Нью-Джерси (2021). Зимой митохондрии эритроцитов птиц производят больше тепла, чем осенью. FASEB J. 35:e21490. дои: 10.1096/fj.202100107R
, Меткалф, Н.Б., Пейдж, Дж.Л., Хакстейбл, А., Маккафферти, Д.Дж., и Доусон, Нью-Джерси (2021). Зимой митохондрии эритроцитов птиц производят больше тепла, чем осенью. FASEB J. 35:e21490. дои: 10.1096/fj.202100107R
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Пенья-Вильялобос И., Нуньес-Вильегас М., Бозинович Ф. и Сабат П. (2014). Метаболические ферменты у сезонно акклиматизированного и акклиматизированного к холоду красношейного воробья, обитающего в чилийской средиземноморской среде. Курс. Зоол. 60, 338–350. doi: 10.1093/czoolo/60.3.338
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Пенья-Вильялобос, И., Вальдес-Ферранти, Ф., и Сабат, П. (2013). Осморегуляторные и метаболические затраты на выделение солей у красношейного воробья Zonotrichia capensis . Комп. Биохим. Физиол. - Мол. интегр. Физиол. 164, 314–318. doi: 10.1016/j.cbpa.2012.10.027
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Квайссер, Н. , Отейза, П.И., Стоппер, Х., Оли, Р.Г., и Шупп, Н. (2011). Альдостерон вызывает окислительный стресс, окислительное повреждение ДНК и активацию NF-κB в клетках почечных канальцев. Мол. Карциног. 50, 123–135. doi: 10.1002/mc.20710
, Отейза, П.И., Стоппер, Х., Оли, Р.Г., и Шупп, Н. (2011). Альдостерон вызывает окислительный стресс, окислительное повреждение ДНК и активацию NF-κB в клетках почечных канальцев. Мол. Карциног. 50, 123–135. doi: 10.1002/mc.20710
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Рахман, И., Коде, А., и Бисвас, С.К. (2006). Анализ для количественного определения уровней глутатиона и глутатиондисульфида с использованием метода ферментативной переработки. Нац. протокол 1, 3159–3165. doi: 10.1038/nprot.2006.378
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Рибейро, Дж. П., Магалхаес, Л. М., Рейс, С., Лима, Дж. Л., и Сегундо, М. А. (2011). Высокая пропускная способность общего количества ионов меди, снижающая антиоксидантную способность биологических образцов, определяемая с помощью проточного инжекторного анализа и методов на основе микропланшетов. Анал. науч. 27, 483–483. doi: 10.2116/analsci. 27.483
27.483
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Райли, Дж.Л., Бакстер-Гилберт, Дж. Х., Гульельмо, К. Г., и Литцгус, Дж. Д. (2016). Сканирование змей для измерения состояния: подтверждение количественного магнитного резонанса. Дж. Герпетол. 50, 627–632. дои: 10.1670/15-113
Полнотекстовая перекрестная ссылка | Академия Google
Сабат, П., Бозинович, Ф., Контрерас-Рамос, К., Несполо, Р.Ф., Ньюсом, С.Д., Киричи, В., и др. (2019). Взаимодействие между температурой окружающей среды и потреблением соли влияет на окислительный статус и иммунные реакции у вездесущего неотропического воробья, красношейного воробья. Комп. Биохим. Физиол. Мол. интегр. Физиол. 234, 50–59. doi: 10.1016/j.cbpa.2019.04.016
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Сабат, П., Гонсалес-Вехарес, С., и Мальдонадо, К. (2009). Диета и засушливость среды обитания влияют на осморегулирующую физиологию: внутривидовое полевое исследование вдоль градиентов окружающей среды у красношейного воробья. Комп. Биохим. Физиол. Мол. интегр. Физиол. 152, 322–326. doi: 10.1016/j.cbpa.2008.11.003
Комп. Биохим. Физиол. Мол. интегр. Физиол. 152, 322–326. doi: 10.1016/j.cbpa.2008.11.003
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Сабат, П., Мальдонадо, К., Ривера-Хутинель, А., и Фарфан, Г. (2004a). Преодоление соли без солевых желез: осморегуляторная пластичность у трех видов прибрежных певчих птиц (печников) рода Cinclodes (Passeriformes: Furnariidae). Дж. Комп. Физиол. В 174, 415–420. doi: 10.1007/s00360-004-0428-422
Полнотекстовая перекрестная ссылка | Академия Google
Сабат П., Сепульведа-Каттан Э. и Мальдонадо К. (2004b). Физиологические и биохимические реакции на пищевой белок у всеядных воробьиных Zonotrichia capensis (Emberizidae). Комп. Биохим. Физиол. Мол. интегр. Физиол. 137, 391–396. doi: 10.1016/j.cbpb.2003.10.021
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Сабат, П., и Мартинес дель Рио, К. (2002). Меж- и внутривидовая изменчивость использования морских пищевых ресурсов тремя видами Cinclodes (Furnariidae, Aves): изотопы углерода и осморегуляторная физиология. Зоология 105, 247–256. дои: 10.1078/0944-2006-00078
Зоология 105, 247–256. дои: 10.1078/0944-2006-00078
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Сабат, П., Нарваэс, К., Пенья-Вильялобос, И., Контрерас, К., Мальдонадо, К., Санчес-Эрнандес, Дж. К., и соавт. (2017). Приспособление к среде обитания соленой воды: метаболические и окислительные реакции на потребление соли у красношейного воробья. Фронт. Физиол. 8:654. doi: 10.3389/fphys.2017.00654
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Сабат П., Ньюсом С. Д., Пиночет С., Несполо Р., Санчес-Эрнандес Дж. К., Мальдонадо К. и др. (2021). Измерения тройного изотопа кислорода (Δ’17O) в воде тела отражают потребление воды, метаболизм и δ18O проглоченной воды у воробьиных. Фронт. Физиол. 12:710026. doi: 10.3389/fphys.2021.710026
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Салин, К., Ауэр, С.К., Рудольф, А.М., Андерсон, Г.Дж., Кэрнс, А. Г., Маллен, В., и соавт. (2015). Люди с более высокой скоростью метаболизма имеют более низкие уровни активных форм кислорода in vivo. Биол. лат. 11:20150538. doi: 10.1098/rsbl.2015.0538
Г., Маллен, В., и соавт. (2015). Люди с более высокой скоростью метаболизма имеют более низкие уровни активных форм кислорода in vivo. Биол. лат. 11:20150538. doi: 10.1098/rsbl.2015.0538
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Шмидт-Нильсен, К.(1997). Физиология животных: адаптация и окружающая среда. Кембридж: Издательство Кембриджского университета.
Академия Google
Смит, Б., и Маккечни, А. Э. (2015). Потоки воды и энергии летом у воробьиных птиц аридной зоны. Ибис 157, 774–786. doi: 10.1111/ibi.12284
Полнотекстовая перекрестная ссылка | Академия Google
Стиер, А., Бизе, П., Шулл, К., Золл, Дж., Сингх, Ф., Джени, Б., и другие. (2013). Птичьи эритроциты имеют функциональные митохондрии, что открывает новые перспективы для птиц как животных моделей в изучении старения. Фронт. Зоол. 10:33. дои: 10.1186/1742-9994-10-33
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Стир, А. , Райхерт, С., Крискуоло, Ф., и Бизе, П. (2015). Эритроциты открывают многообещающие возможности для лонгитюдных исследований старения в лабораторных, немодельных и диких животных. Экспл. Геронтол. 71, 118–134. doi: 10.1016/j.exger.2015.09.001
, Райхерт, С., Крискуоло, Ф., и Бизе, П. (2015). Эритроциты открывают многообещающие возможности для лонгитюдных исследований старения в лабораторных, немодельных и диких животных. Экспл. Геронтол. 71, 118–134. doi: 10.1016/j.exger.2015.09.001
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Стир, А., Роместинг, К., Schull, Q., Lefol, E., Robin, J.P., Roussel, D., et al. (2017). Как измерить митохондриальную функцию у птиц с помощью эритроцитов: тематическое исследование королевского пингвина и перспективы экологии и эволюции. Методы Экол. Эвол. 8, 1172–1182. дои: 10.1111/2041-210X.12724
Полнотекстовая перекрестная ссылка | Академия Google
Стерки П.Д. и Гримингер П. (1986). «Жидкости организма: кровь», в Птичья физиология , изд. PD Sturkie (Нью-Йорк, штат Нью-Йорк: Springer), doi: 10.1007/978-1-4612-4862-0_5
Полнотекстовая перекрестная ссылка | Академия Google
Тапиа-Монсалве, Р. , Ньюсом, С. Д., Санчес-Эрнандес, Дж. К., Бозинович, Ф., Несполо, Р., и Сабат, П. (2018). Наземные птицы в прибрежной среде: скорость метаболизма и окислительный статус варьируются в зависимости от использования морских ресурсов. Экология 188, 65–73. doi: 10.1007/s00442-018-4181-8
, Ньюсом, С. Д., Санчес-Эрнандес, Дж. К., Бозинович, Ф., Несполо, Р., и Сабат, П. (2018). Наземные птицы в прибрежной среде: скорость метаболизма и окислительный статус варьируются в зависимости от использования морских ресурсов. Экология 188, 65–73. doi: 10.1007/s00442-018-4181-8
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Тилеман, Б.И., Уильямс, Дж.Б., Бущур, М.Е., и Браун, Ч.Р. (2003). Фенотипическая изменчивость жаворонков по градиенту засушливости: пустынные птицы более гибкие? Экология 84, 1800–1815.
Академия Google
Тумминелло, Р. А., и Фуллер-Эспи, С. Л. (2013). Тепловой стресс индуцирует продукцию АФК и фосфорилирование гистонов в целомоцитах Eisenia hortensis . ISJ 10, 50–57.
Академия Google
Веландо, А., Ногера, Дж. К., да Силва, А., и Ким, С.Ю. (2019). Окислительно-восстановительная регуляция и компромиссы в жизненном цикле: удаление митохондриальных АФК улучшает рост дикой птицы. Науч. Респ. 9:2203. doi: 10.1038/s41598-019-38535-5
Науч. Респ. 9:2203. doi: 10.1038/s41598-019-38535-5
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Везина, Ф., и Уильямс, Т. (2005). Взаимодействие между массой органов и активностью цитратсинтазы как показатель максимальной окислительной способности тканей при разведении европейских скворцов: последствия для скорости обмена веществ и соотношения массы органов. Функц. Экол. 19, 119–128.
Академия Google
Вальсберг, Г. Э., и Вольф, Б. О. (1995). Изменение коэффициента респирометрии птиц и последствия для непрямой калориметрии с использованием измерений производства углекислого газа. Дж. Экспл. биол. 198, 213–219. doi: 10.1242/jeb.198.1.213
Полнотекстовая перекрестная ссылка | Академия Google
Walther, G.R., Post, E., Convey, P., Menzel, A., Parmesan, C., Beebee, T.J., et al. (2002). Экологические реакции на недавнее изменение климата. Природа 416, 389–395. дои: 10.1038/416389a
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Уайтман, Дж. П., Шарп, З. Д., Герсон, А. Р., и Ньюсом, С. Д. (2019). Связь значений Δ17O в воде тела животного с поступлением экзогенной воды и метаболизмом. BioScience 69, 658–668. doi: 10.1093/biosci/biz055
П., Шарп, З. Д., Герсон, А. Р., и Ньюсом, С. Д. (2019). Связь значений Δ17O в воде тела животного с поступлением экзогенной воды и метаболизмом. BioScience 69, 658–668. doi: 10.1093/biosci/biz055
Полнотекстовая перекрестная ссылка | Академия Google
Уизерс, ПК (1977). Дыхание, обмен веществ и теплообмен эутермических и вялотекущих бедвилл и колибри. Физиол. Зоол. 50, 43–52. doi: 10.1086/physzool.50.1.30155714
Полнотекстовая перекрестная ссылка | Академия Google
Памяти профессора Эфраима Мориса Воробья | J. Теплопередача
В четверг, 1 августа 2019 г., мы были глубоко опечалены известием о том, что профессор Эфраим М. Воробей скончался в возрасте 91 года. Профессор Эфраим М. Воробей родился 27 мая 1928 г. и благодаря своей обширной профессиональной карьере стал одним из самых известных и уважаемых исследователей в области теплопередачи.Он получил степень бакалавра наук. и М.С. в 1948 и 1949 годах, соответственно, из Массачусетского технологического института (MIT). Затем он переехал в Гарвардский университет, чтобы получить степень магистра и доктора философии. в 1950 и 1952 годах соответственно. Его докторская степень Тема диссертации: «Свободная конвекция с переменными свойствами и переменной температурой стенок» под руководством профессора Говарда Уилсона Эммонса.
в 1948 и 1949 годах, соответственно, из Массачусетского технологического института (MIT). Затем он переехал в Гарвардский университет, чтобы получить степень магистра и доктора философии. в 1950 и 1952 годах соответственно. Его докторская степень Тема диссертации: «Свободная конвекция с переменными свойствами и переменной температурой стенок» под руководством профессора Говарда Уилсона Эммонса.
После получения докторской степени. в 1952 году профессор Эфраим М.Воробей присоединился к Raytheon, где он работал над различными проблемами теплопередачи, такими как радар, охлаждение электроники и первая микроволновая печь. Позже он присоединился к Национальному консультативному комитету по аэронавтике (NACA) (теперь Национальное управление по аэронавтике и исследованию космического пространства (НАСА)), Лаборатории летных двигателей Льюиса, Кливленд, Огайо, где он работал над гидромеханикой и теплопередачей. У него было много публикаций о его работе в НАСА, таких как две статьи [1,2] в первом выпуске ASME Journal of Heat Transfer (JHT), который был опубликован в феврале 1959 года с 15 статьями, двумя статьями [3,4 ] во втором выпуске JHT (май 1959 г. ), одна статья [5] в третьем выпуске JHT (август 1959 г.) и две статьи [6,7] в четвертом выпуске JHT (ноябрь 1959 г.).
), одна статья [5] в третьем выпуске JHT (август 1959 г.) и две статьи [6,7] в четвертом выпуске JHT (ноябрь 1959 г.).
В 1959 году он начал свою карьеру в Лаборатории термодинамики и теплопередачи (THTL), факультет машиностроения, Миннесотский университет, Миннеаполис, Миннесота. THTL была основана профессором Эрнстом Р. Г. Эккертом в 1950 году. В период с 1968 по 1980 год профессор Эфраим М. Воробей был руководителем программы по гидромеханике. В период 1986–1988 гг. профессор Эфраим М. Воробей работал в Национальном научном фонде (NSF), сначала в качестве руководителя программы теплопередачи, а затем в качестве директора отдела CBET в инженерном управлении.В NSF он подчеркнул необходимость сосредоточиться на практических проблемах и важных фундаментальных вопросах. На момент своей смерти он был как самым старым преподавателем, так и самым старым на факультете машиностроения Миннесотского университета, Миннеаполис, Миннесота.
За свою академическую жизнь профессор Воробей подготовил более 250 магистрантов и более 100 докторантов. Многие из его предыдущих докторских диссертаций.Аспиранты Д. стали профессорами известных университетов мира. Профессор Воробей опубликовал множество статей, отчетов и книг по гидромеханике и теплопередаче, таких как Refs. [8] и [9]. Его статья в журнале ASME Journal of Heat Transfer (JHT), которая была опубликована в мае 1977 г., о «Полностью развитом потоке и теплопередаче в каналах, имеющих продольно-периодические изменения площади поперечного сечения» [10], была его наиболее цитируемой статьей. Он был пионером в исследованиях во многих важных областях теплопередачи, и его обширный вклад послужил огромным стимулом для исследователей и инженеров по всему миру.
В мае 1988 года 60-летие Эфа было отмечено статьей, опубликованной в International Journal of Heat and Mass Transfer [11]. После своего 60-летия в 1988 году и до своей смерти в 2019 году Эф не проявлял никаких признаков замедления темпов роста и, в своей типичной манере, очень активно преследовал свои исследовательские интересы. Профессор Эфраим М. Воробей был очень активен до последнего года своей жизни. Например, у него были эти публикации в 2019 году [12–18].
После своего 60-летия в 1988 году и до своей смерти в 2019 году Эф не проявлял никаких признаков замедления темпов роста и, в своей типичной манере, очень активно преследовал свои исследовательские интересы. Профессор Эфраим М. Воробей был очень активен до последнего года своей жизни. Например, у него были эти публикации в 2019 году [12–18].
Во время своей академической карьеры, после профессора У. Х. Гидта в 1972 году, профессор Эфраим М. Спарроу работал редактором журнала ASME Journal of Heat Transfer до 1980 года, когда эту должность сменил профессор К. Т. Ян.
В 1979 году он вместе с профессором В. Дж. Минковичем основал журнал под названием Numerical Heat Transfer , чтобы служить форумом для распространения идей и исследований в области численного теплообмена и вычислительной гидродинамики (CFD).Много раз был приглашенным редактором. Например, он был приглашенным редактором специального выпуска, связанного с биомедициной, в International Journal of Heat and Mass Transfer с профессором Джоном П. Абрахамом, Университет Сент-Томас, в ноябре 2008 г. [19]. Он был соавтором с профессорами Дж. П. Абрахамом и Р. Д. Ловиком в статье о «Нестационарном трехмерном гидромеханическом анализе кровотока в артериях, суженных и свободных от бляшек» в специальном выпуске, связанном с биомедициной [20].Кроме того, том 50 « Достижения в области теплопередачи» редактировали Эфраим М. Воробей, Джон П. Абрахам и Джон М. Горман. 1 В томе шесть глав. Профессор Эфраим М. Воробей был соавтором первой главы об «Улучшении теплопередачи струйного удара посредством переключения оси струи» [21].
Например, он был приглашенным редактором специального выпуска, связанного с биомедициной, в International Journal of Heat and Mass Transfer с профессором Джоном П. Абрахамом, Университет Сент-Томас, в ноябре 2008 г. [19]. Он был соавтором с профессорами Дж. П. Абрахамом и Р. Д. Ловиком в статье о «Нестационарном трехмерном гидромеханическом анализе кровотока в артериях, суженных и свободных от бляшек» в специальном выпуске, связанном с биомедициной [20].Кроме того, том 50 « Достижения в области теплопередачи» редактировали Эфраим М. Воробей, Джон П. Абрахам и Джон М. Горман. 1 В томе шесть глав. Профессор Эфраим М. Воробей был соавтором первой главы об «Улучшении теплопередачи струйного удара посредством переключения оси струи» [21].
Кроме того, вклад профессора Эфраима М. Воробья охватывает многие области и был отмечен многочисленными почестями и наградами.Среди них: Мемориальная премия Макса Якоба в 1976 г. , член Национальной инженерной академии в 1986 г., премия Мони А. Ферст в 1993 г., член Американского общества инженеров-механиков (ASME) и медаль ASME Ральфа Коутса Роу.
, член Национальной инженерной академии в 1986 г., премия Мони А. Ферст в 1993 г., член Американского общества инженеров-механиков (ASME) и медаль ASME Ральфа Коутса Роу.
В июле 2020 года будет открыт Мемориальный трек профессора Эфраима Воробья, посвященный его жизни и исследовательскому вкладу. Этот специальный симпозиум будет организован Отделом теплопередачи (HTD) в его память.Участие в этом симпозиуме принимается из всех соответствующих технических тем, связанных с личными воспоминаниями о трех программах: ICNMM 2020, SHTC 2020 и FEDSM 2020 в Розен Шингл Крик, Орландо, Флорида, 12–15 июля 2020 г. 2
Трудно подобрать слова, чтобы выразить свои чувства в этот печальный момент, но, несомненно, профессиональное сообщество запомнит профессора Ефрема М. Воробья как прекрасного и плодовитого ученого, педагога, наставника и, прежде всего, приятного и честного человека, и очень уважаемый коллега с выдающимся характером и этикой.