
Лесная соня
Лесная соня Dryomys nitedula (Pallas, 1778) Отряд Грызуны Rodentia Семейство Соневые Myoxidae
Cтатус. Категория 4 вид, неопределенный по статусу.
Статус в России и сопредельных регионах. Вид занесен в Красные книги Липецкой, Орловской, Курской и Тамбовской обл.
Статус международный. Вид внесен в Красный список МСОП, охраняется Бернской конвенцией.
Распространение.
Широко распространен по зоне лесов от Европы до Казахстана и Центральной Азии. В Воронежской обл. обитает повсеместно от крупных лесных массивов до лесных полос и небольших байрачных лесов. Отмечены встречи у северо-западной окраины Воронежа, в ХГПЗ, ВГПБЗ, в Шиповом лесу, Хреновском лесу.
Описание.
Зверек средней величины с длиной тела до 110 мм и длинным хвостом до 113 мм. Хвост хорошо опушен и слегка уплощен сверху вниз. Передние конечности четырехпалые, задние пятипалые, с острыми сильно изогнутыми когтями. Довольно пушистый мех, на верхней стороне тела серовато-охристый, на боках серее, а на горле, груди и брюшке серовато-желтый. Хвост грязно-серый. Между носом и основанием уха проходит черная полоса, едва заходящая за ухо.
Особенности биологии и экологии.
Придерживается широколиственных и смешанных лесов с хорошо развитым подлеском. Ведет древесный образ жизни. Хорошо лазает по деревьям. Под убежища использует неглубокие норы, пустоты под корнями, дупла, искусственные птичьи гнездовья, старые сорочьи гнезда, реже строит сама шарообразные гнезда из прутьев на кустарниках и деревьях. Активна по ночам. На зиму впадает в спячку. Питается плодами, почками, корой деревьев, насекомыми. При случае съедает яйца птиц, птенцов и мелких зверьков. Иногда поселяется в садах, где может вредить плодовым культурам.
Численность и тенденции ее изменения. Везде низкая. За последние 20 лет наблюдений в окр. биоцентра ВГУ на 9 800 ловушко-ночей была отловлена всего 1 лесная соня. Однако в разные годы при ежегодном осмотре синичников были обнаружены 8 взрослых сонь и 2 выводка по 6 молодых зверьков этого вида, устраивавшихся там на дневку.
Везде низкая. За последние 20 лет наблюдений в окр. биоцентра ВГУ на 9 800 ловушко-ночей была отловлена всего 1 лесная соня. Однако в разные годы при ежегодном осмотре синичников были обнаружены 8 взрослых сонь и 2 выводка по 6 молодых зверьков этого вида, устраивавшихся там на дневку.
Лимитирующие факторы. Нарушение естественных лесных местообитаний, слабое развитие подлеска, сокращение числа старых дуплистых деревьев, плохое плодоношение деревьев и кустарников.
Принятые и необходимые меры охраны. Охраняется в ВГПБЗ и ХГПЗ. Требуется сохранение естественных лесных биоценозов.
Источники информации: Красная книга Воронежской области 1. Барабаш-Никифоров, 1957. 2 . Млекопитающие фауны СССР, 1963. 3. Айрапетьянц, 1983. 4. Громов, 1995. 5 . Климов А. С ., 1996б. 6. Наземные звери... 2002. Составитель: А. С. Климов; фото: А. Д. Нумеров.
AOF | 10.01.2016 12:02:31
Лесная соня — описание, фотографии, отзывы, места — Млекопитающие Удмуртии
Наименование: ЛЕСНАЯ СОНЯ — Dryomys nitedula (Pallas, 1779)
Отряд: Грызуны.
Семейство: Соневые.
Природоохранный статус: 3 категория. Вид занесен в Красную книгу Республики Татарстан.
Краткое описание: Размеры средние, длина тела до 11 см, примерно такой же величины хвост. Задние конечности заметно больше передних. Уши закругленные, покрыты негустыми волосами. Окраска верха тела серовато-охристая. Хвост сверху темнее и обычно серее спины, нередко с узкой и нерезкой белой оторочкой, снизу беловатый. От носа к основанию уха проходит черная полоса. От внешне сходной садовой сони отличается тем, что эта полоса не заходит за ухо.
Распространение: Современный ареал вида охватывает Центральную и Восточную Европу, Малую и Переднюю Азию, Кавказ и горные системы Средней и Центральной Азии. По территории Удмуртии проходит северная граница видового ареала.
По территории Удмуртии проходит северная граница видового ареала.
Экология: Населяет самые разнообразные лесные биотопы, включая искусственные лесополосы и кустарниковые заросли в горах. В условиях региона отмечена определенная приуроченность вида к пойменным лесам. Селится в дуплах деревьев, иногда в шарообразных гнездах на ветвях деревьев и кустарников. Охотно занимает брошенные гнезда птиц и различные искусственные гнездовья. Для зимней спячки переселяется в подземные норы, которые устраивает обычно под корнями и кучами хвороста. Размножается обычно один раз в год, в помете 2 — 9 детенышей. Основными естественными врагами лесной сони являются совы и мелкие куньи, в антропогенных ландшафтах также домашняя кошка.
Современное состояние: В республике лесная соня спорадически встречается в Сюмсинском, Вавожском и Кизнерском районах.
Лимитирующие факторы: Вырубка спелых и старовозрастных лесов.
Меры охраны: Выяснение распространения в пределах Удмуртии, организация заказников в местах обитания.
Источники информации: 1. Красная..., 2006; 2. Айрапетьянц, 1983; 3. Громов, Ербаева, 1995; 4. Россолимо и др., 2001; 5. Попов, 1972; 6. Капитонов и др., 1997.
Автор-составитель: Капитонов В.И.
Рисунок: Иванов В.В.
Лесная соня. В Красной книге России и под охраной | Животные мира|Редкие животные
Здравствуйте, друзья!
Один из самых древних грызунов нашей планеты – Лесная соня. Зверек немного видом похож на маленькую белку – у него такой же большой и пушистый хвост.
Описание и места обитания Лесной сони
Окрас у Лесной сони отличается в зависимости от региона обитания. В длину эти грызуны вырастают не больше двенадцати сантиметров, а их хвост имеет примерно такую же длину, как и тело. Он не только им служит как балансир для лазанья по деревьям, но и как у кошек показывает их настроение. Интересный факт – на задних лапках сони 5 пальцев, а на передних их только четыре.
Он не только им служит как балансир для лазанья по деревьям, но и как у кошек показывает их настроение. Интересный факт – на задних лапках сони 5 пальцев, а на передних их только четыре.
Лесные сони проживают в лесных зонах Центральной Азии, Казахстана и стран Европы, северной части Африки, Китая и Японии. Также они встречаются в Малой Азии и на Алтае. Этот милый зверек так же проживает в средней полосе и на западе европейской части России.
Повадки и рацион Лесной сони
Основным местом проживания лесной сони являются лиственные леса и густые заросли. Там они могут обустроить себе дом. В основном это заброшенные гнезда птиц или дупла деревьев. Но если соня не найдет себе приют, она запросто может влезть в заселенное птицами гнездо и выжить оттуда хозяев.
Но при нужде они могут и сами построить себе жилище где-нибудь в глухих зарослях, чтобы защититься от хищников.
Питаются сони в основном растительной пищей, но иногда разбавляют свой рацион насекомыми. Но запросто может перекусить и некрупными птичьими яйцами.
С приближением холодов лесные сони делают себе норы для спячки на глубину порядка тридцати сантиметров. Запасаются пищей и с наступлением морозов впадают в спячку. Если во время зимы наступит оттепель, соня может проснуться от потепления, перекусить своими запасами и решив, что нормальное тепло еще не пришло, уснуть дальше.
Размножение Лесной сони
Сразу же после своего пробуждения начинается период их размножения, и грызуны активно начинают искать партнеров. Они не строят семейных отношений и после спаривания с детьми остается только мама. Беременность у сони продолжается меньше месяца, а когда мама растит детенышей, она отлучается от гнезда исключительно перекусить.
На свет появляется не больше восьми малышей. В возрасте примерно полтора месяца детеныши становятся вполне самостоятельными, но, если в запасах достаточно пищи, они могут остаться жить группой.
Статус вида Лесная соня
Лесные сони легко приручаются, но только во младенчестве.
Основной враг популяции лесной сони – это совы.
Несмотря на то, что лесные сони достаточно распространены, они занесены в Красную книгу в части регионов России и находятся под охраной МСОП.
Понравилась статья? Нажимайте палец вверх, оставляйте комментарии и подписывайтесь на канал, чтобы не пропускать свежие публикации.
Лучшие статьи (по мнению читателей) канала о редких животных вы можете посмотреть по ЭТОЙ ССЫЛКЕ
где обитает, описание, фото, жизнь в природе
Сони (Gliridae) – небольшие грызуны с мягким пушистым мехом, внешне напоминающие белок. Представители этого семейства распространены в Европе, Африке и южной половине Азии. Четыре вида сонь живут в России, и об одном из этих видов, лесной соне, расскажем в этой статье.
Где обитает лесная соня?
Лесная соня (Dryomys nitedula) встречается на Балканском полуострове, в Центральной и Восточной Европе, на юге Европейской части России, в Средней и Центральной Азии, на Кавказе, в горах западной Монголии и в южной и восточной частях Казахстана.
Места обитания лесной сони связаны с древесной и кустарниковой лиственной растительностью. Она встречаются в густых лесах, обычно лиственных и смешанных, а также в зарослях кустарника. В горы поднимаются до высоты 3500 м. Иногда держится по каменистым осыпям, поросшим кустарником, селится на лесных опушках, каменистых лугах, в культурных садах населенных пунктов, заселяет просеки.
Описание и фото лесной сони
Лесная соня – небольшой красивый зверек с большими глазами, длинным пушистым хвостом и закругленными ушками.
Лапки сони имеют признаки приспособления к древесному образу жизни. Третий палец задних конечностей самый длинный.
Окраска меха спины варьирует от светлой серовато-охристо-бурой до более темной серо-коричневой. Средняя часть носа и лба черноватые. Вокруг глаз и под ушами темные, черноватые пятна, значительно не доходящие до конца носа.
Мех молодых особей серее меха взрослых, короче и ровнее его.
Хвост уплощенный с густыми, довольно длинными, однообразно окрашенными в серый цвет, как бы расчесанными на стороны волосами.
На фото лесная соня демонстрирует свой пушистый хвост.
Уши покрыты короткими, относительно редкими волосами.
Череп заметно приподнят по профилю, особенно в затылочной части; носовая часть относительно узкая, а затылочная – закругленная. Межглазничная часть не уплощенная.
Длина тела лесной сони составляет в среднем 8-13 см, длина хвоста – 6-13 см, масса тела – 18-34 г. Внешне лесная соня очень похожа на соню садовую (Eliomys quercinus). О других видах сонь можно узнать здесь.
Образ жизни и поведение лесной сони
Лесная соня – очень подвижный грызун, прекрасно бегающий по земле и лазающий по стволам деревьев и веткам, а также по скалистым выступам. В случае опасности зверек иногда спрыгивает на землю с довольно большой высоты. Активными сони бывают чаще в ночное время, а днем обычно спят в норах или гнездах.
Гнезда лесные сони строят сами или используют птичьи. Гнезда располагаются как на ветках деревьев и в дуплах, так и на кустарниках, причем на разной высоте от земли. В горных местностях они нередко занимают норы других лесных грызунов или устраивают свои.
Гнезда у зверька могут быть временные и постоянные. Временные гнезда – непрочные сооружения; гораздо больше усилий грызуны вкладывают в строительство гнезд, в котором будет жить потомство. Такие гнезда обычно находятся на высоте от 1 до 7 м над уровнем земли и имеют диаметр от 15 до 25 см. Строятся они из листьев и веток и выстилаются кусочками коры или мха.
В зимний период сони, как правило, впадают в спячку. В северных районах их распространения, например в Европе, лесные сони зимуют с октября по апрель. В южных частях ареала в теплые зимы спячка бывает непродолжительной, и по-видимому, неглубокой.
Во время зимней спячки соня садится на задние лапы, сворачивается в клубок, обвивает хвост вокруг тела и прижимает лапки к щекам. Иногда зверек просыпается, чтобы поесть что-нибудь из того, что заготовил заранее.
Иногда зверек просыпается, чтобы поесть что-нибудь из того, что заготовил заранее.
Лесные сони – зверьки территориальные. Размеры их индивидуальных участков колеблются от 65 до 100 м в диаметре.
Эти зверьки издают различные звуки. Наиболее заметным из них является тонкий, мелодичный писк, который, по-видимому, является сигналом тревоги.
Питание
Лесные сони всеядны. Преимущественно они питаются плодами, ягодами и семенами деревьев и кустарников, среди которых живут, при этом не избегают и животной пищи, в большом количестве поедая насекомых. При случае не откажется перекусить и яйцами мелких птиц.
Размножение
О брачном поведении этого вида известно не так уж много. Сезон размножения лесной сони варьирует по всему ареалу. В Европе брачный сезон длится с мая по август, при этом каждый год появляется только один помет. Кое-где, например в Израиле, брачный сезон приходится на март – декабрь, а каждая самка в среднем рожает 2-3 раза в год.
Срок беременности составляет от 21 до 30 дней. В помете бывает 2-7 детенышей, чаще всего 3-4. Новорожденные весят не более 2-3 граммов и совершенно беспомощны. Глаза они открывают примерно на 16-17 день. Пока детеныши не станут самостоятельными, до достижения ими возраста 4-5 недель, мать кормит их, ухаживает и защищает от опасностей. Принимают ли самцы какое-то участие в заботе о потомстве, ученым не известно.
Лесная соня занесена в региональные Красные книги Орловской, Курской, Липецкой и Тамбовской областей.
Считается, что в природных условиях продолжительность жизни лесной сони составляет в среднем 5,5 лет.
Вконтакте
Одноклассники
Мой мир
Лесная соня (краткое описание для детей)
Лесная соня – маленький дикий грызун, похожий на хомяка с длинным пушистым хостом. Длина тела у сони около 10 сантиметров, еще 10 сантиметров добавляет ей хвост. Шерстка у сони очень мягкая и шелковистая и имеет серовато-бежевый окрас. От носа к ушам по обе стороны мордочки идет темная полоса, по которой лесную соню легко отличить от других грызунов. Благодаря цепким коготкам на лапах, соня хорошо лазает по деревьям.
От носа к ушам по обе стороны мордочки идет темная полоса, по которой лесную соню легко отличить от других грызунов. Благодаря цепким коготкам на лапах, соня хорошо лазает по деревьям.
На мордочке у лесной сони два больших черных глаза, круглые уши, нос и усы. Усы лесной сони называются вибриссами. Они очень чувствительные и являются дополнительным органом чувств. Даже в темноте соня может ориентироваться в пространстве, улавливая колебания воздуха своими вибриссами.
Мех на хвосте у сони заметно длиннее, чем на теле. Хвост играет важную роль в жизни лесной сони. Также как и у кошек, хвост помогает ей сохранять равновесие при прыжках. Когда лесная соня испугана или злится, то мех на хвосте распушается в разные стороны и хвост выглядит еще больше.
Лесные сони очень чистоплотные. Они много времени тратят на вычесывание своей шерсти и содержат свое жилище в чистоте.
Уже из названия понятно, что проживают лесные сони в лесах. Чаще всего они селятся там, где растут дубы, потому то они питаются желудями. Кроме того, лесные сони могут лакомиться буковыми орешками, яблочными семечками и мякотью лесных плодов. Иногда сони ловят и поедают насекомых.
Жилище лесная соня строит в укромном месте из прутиков, травы, сухих листьев. Внутренние стены соня утепляет пухом и сухими травинками. Но не всегда она сооружает свой собственный дом, иногда соня занимает дупла деревьев или пустые птичьи гнезда. К зиме лесная соня начинает собирать у себя в домике съестные запасы.
Зимой лесная соня впадает в спячку. Время от времени она просыпается чтобы перекусить, но большую часть холодов соня проводит во сне.
В некоторых регионах России лесная соня занесена в Красную книгу.
Соня лесная — Dryomys nitedula
Тип Хордовые Chordata
Подтип Позвоночные Vertebrata
Класс Млекопитающие Mammalia
Отряд Грызуны Rodentia
Семейство Соневые Myoxidae
СОНЯ ЛЕСНАЯ Dryomys nitedula (Pallas, 1778) BĂPMAH ÇЫBPABÇИ
Статус. IV категория. Вид с неопределённым статусом. Внесён в Красную книгу МСОП.
IV категория. Вид с неопределённым статусом. Внесён в Красную книгу МСОП.
Описание. Длина тела достигает 9-11 см, вес — 25-45 г, хвоста — 7-9,6 см. Мех густой, хорошо выражена ость. Окраска светло-коричневая, сверху рыжевато- или серовато-охристая, снизу светлая. От уха к глазу через нос проходит чёрная полоса. Хвост покрыт длинной густой шерстью, его кончик светлый. Лазает по деревьям, собирая ягоды, семена, листья, почки и плоды, ест мелких беспозвоночных, иногда птичьи яйца. Использует старые дуплистые деревья, птичьи гнёзда, строит гнёзда на ветвях деревьев. Зимние гнёзда устраивает под корнями деревьев. Активен в сумерках и ночью. На зиму впадает в спячку, делает к зиме запасы, которыми пользуется, когда просыпается. Приступает к размножению в мае. Даёт один приплод по 3-6 детёнышей, которые переходят к самостоятельному образу жизни в возрасте 2-2,5 месяцев. Половозрелость наступает после первой зимовки. Продолжительность жизни около — 3-4 лет. Ареал — Центральная, Юго-Восточая и Восточная Европа, Кавказ, горы Средней Азии, Алтай. В Чувашии обитает в Присурье, Предволжье и Центре. Внесён в Красные книги Нижегородской (категория Д), Ульяновской (III категория) областей, республик Татарстан (III категория), Мордовия (Приложение — Аннотированный перечень таксонов животных, нуждающихся в особом внимании к их состоянию в природной среде, 2008).
Места обитания. Лиственные и смешанные леса с преобладанием дуба и липы, где хорошо развит подлесок. Встречается также в плодовых садах.
Численность и тенденции её изменения. Малочислен. Тенденции изменения численности не изучены.
Основные лимитирующие факторы. Уничтожение дуплистых старовозрастных деревьев из-за вырубок, проведения лесохозяйственных мероприятий. Нерегулярные урожаи лещины, фруктовых деревьев, неблагоприятные условия зимовки.
Разведение. Мероприятия по разведению в республике не проводились. Биотехнические мероприятия для восстановления численности вида не разработаны.
Принятые меры охраны. Местообитания охраняются на территории национального парка «Чăваш вăрманĕ», Алатырского участка государственного природного заповедника «Присурский».
Необходимые меры охраны. Выявление и сохранение местообитаний. Организация особо охраняемых природных территорий. Сохранение в лесах дуплистых старовозрастных деревьев. Развешивание в лесах дуплянок с расширенными летками. Мониторинг численности вида в природных условиях.
Источники информации: Гольцмайер, 1934; Львова, 1952; Пузанов и др., 1955; Попов, 1960; Олигер и др., 1966; Андроников, Фаршатов, 1971 а; Олигер, 1971; Соколов, 1977; Игнатьев, Павлов, 1993; Игнатьев, 1995; данные составителей.
Составители: Плечова З.Н., Димитриев А.В.
Фотогалерея:
Соня (Gliridae или Myoxidae): интересные факты, фото, виды
Соневые – это семейство млекопитающих из отряда грызунов. Их размеры так малы, что сони способны поместиться на ладони человека. Кроме того, эти крошки отличаются длинным пушистым хвостом, похожим на беличий.
Описание грызуна
- Реклама -
Внешне сони напоминают мышей или белок. Длина тела у них 8-20 см. Хвост в длину от 4 до 17 см, густо опушенный у большинства видов, только у селевинии и мышевидных сонь – полуголый. Уши округлой формы, без кисточек. Лапы короткие, с хорошо развитыми острыми коготками. Мех густой и мягкий. Спинка окрашена от серого до охристо-бурого цвета.
Особенности питания сони
Сони питаются и растительными, и животными кормами. В качестве растительной пищи употребляют семена и плоды деревьев и кустарников (желуди, липовые орешки, буковые орешки, яблоки). Среди животных кормов отдают предпочтение насекомым, реже кушают яйца и маленьких птенцов. Состав рациона для каждого вида тесно с сезоном и местами обитания.
Распространение сони
- Реклама -
Ареал распространения видов сони включает Северную Африку, Европу до Южной Скандинавии и Малую Азию до Алтая, северо-запада Китая и Японии.
Распространенные виды сони
Различают множество видов сонь, однако внешне они выглядят практически одинаково и отличаются, прежде всего, регионом обитания.
Ангольская соня (Graphiurus angolensis)
Вид является эндемиком Анголы, где обитает в тропических и субтропических сухих лесах.
Соня Кристи (Graphiurus christyi)
Распространена в Камеруне и Конго. Живет в субтропических и тропических влажных низменных лесах.
Толстохвостая соня (Graphiurus crassicaudatus)
Ареал обитания вида включает Камерун, Кот-д’Ивуар, Гану, Либерию, Нигерию и Того, Гвинею и Сьерра-Леоне. Встречается в субтропических и тропических влажных низменных лесах.
Соня Джонстона (Graphiurus johnstoni)
Обитает во влажных саваннах Малави, Замбии и Зимбабве.
Соня Хьюта (Graphiurus hueti)
Вид распространен в Камеруне, Гане, Либерии, Нигерии, Сьерра-Леоне и Габоне. Обитает в тропических и субтропических влажных низменных лесах.
Соня Келлена (Graphiurus kelleni)
Регион распространения включает тропические и субтропические сухие леса, сухие и влажные саванны Анголы, Буркина-Фасо, Камеруна, Центрально-Африканской республики, Конго, Гамбии, Кении, Мали, Нигерии, Нигера, Сенегала, Танзании и Уганды.
Западноафриканская соня (Graphiurus lorraineus)
Встречается в Камеруне, Конго, Гане, Гвинее-Бисау, Либерии, Нигерии, Сьерра-Леоне, Танзании, Уганде. Средой обитания являются тропические и субтропические влажные леса, саванны и плантации.
Короткоухая соня (Graphiurus microtis)
Обитает в таких странах, как Ангола, Ботсвана, Эритрея, Эфиопия, Кения, Лесото, Малави, Мозамбик, Намибия, Южная Африка, Судан, Свазиленд, Танзания, Замбия и Зимбабве.
Соня Монарда (Graphiurus monardi)
Распространена в Анголе, Конго и Замбии, где живет во влажных саваннах.
Саванная соня (Graphiurus murinus)
Вид является эндемиком Южной Африки.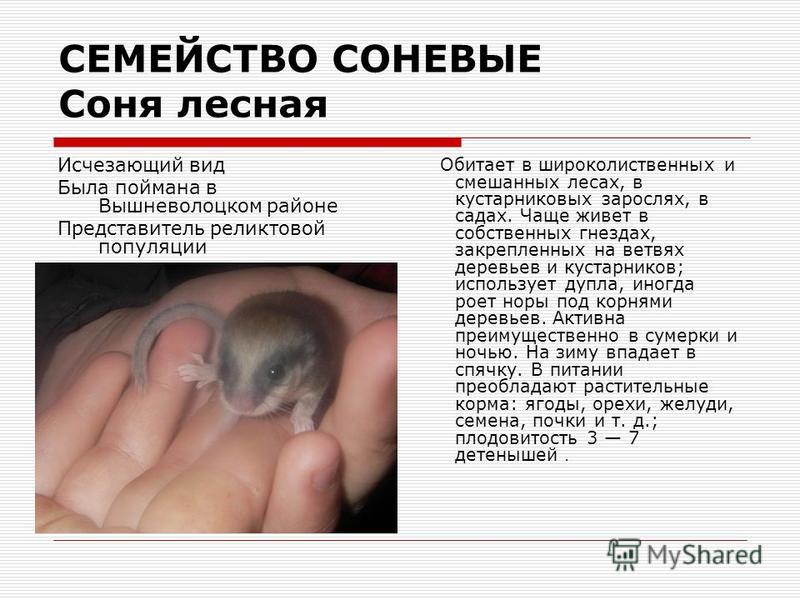 Обитает в субтропических или тропических влажных лесах, неподалеку от рек.
Обитает в субтропических или тропических влажных лесах, неподалеку от рек.
Очковая или южноафриканская соня (Graphiurus ocularis)
Хвост пушистый, напоминающий беличий, с белыми волосками, мордочка короткая, уши маленькие, мех мягкий. На мордочке расположены белые и серые крапинки, вокруг глаз заметны темные круги. Щеки, животик и лапы сверху белого цвета.
Обитают на западе Южной Африки, в скалистых местностях.
Скальная соня (Graphiurus rupicola)
Житель Намибии, Южной Африки и Анголы. Обитает в скалистых местах.
Плоскоголовая соня (Graphiurus platyops)
Встречается в таких странах, как Ангола, Ботсвана, Мозамбик, Свазиленд, Замбия и Зимбабве.
Гвинейская соня (Graphiurus surdus)
Распространена в тропических и субтропических влажных низменных лесах Камеруна, Конго, Экваториальной Гвинеи и Габона.
Анатолийская или пушистая соня (Dryomys laniger)
Вид является эндемиком Турции.
Белуджистанская соня (Dryomys niethammeri)
Встречается исключительно в Пакистане.
Лесная соня (Dryomys nitedula)
Длина тела до 12 см, хвост в длину до 10 см. Вес находится в пределах от 30 до 40 г. Мех серовато-охристый, изменчив. Например, в европейской части России с севера на юг он светлеет. В Закавказье, Апеннинах, Альпах и на Балканском полуострове лесные сони серого цвета. Животик от сероватого до белого цвета. Хвост пушистый, тёмно-серый. От носа к уху проходит темная полоска.
Вид распространен в Европе, Закавказье и Азии в различных типах лесов.
Сычуаньская соня (Chaetocauda sichuanensis)
Длина тела около 9 см, длина хвоста 9-10 см. Масса от 24,5 до 36,0 г.
Вид был обнаружен в смешанных лесах на севере Сычуаня, в Китае. С 2004 года внесен в Красную книгу как вид, находящийся в опасности.
Чернохвостая соня (Eliomys melanurus)
Длина тела 10-18 см, длина хвоста 9-14 см, масса 45-120 г. Спинка бурого цвета, животик белесый. На глазах расположена темная «маска». Хвост пушистый, верхушка черная.
На глазах расположена темная «маска». Хвост пушистый, верхушка черная.
Вид распространен от северо-востока Ливии до запада Аравийского полуострова, и вплоть до южной Турции.
Садовая соня (Eliomys quercinus)
Длина тела от 11 до 16 см, длина хвоста от 9 до 14 см, масса составляет 60-140 г. Мордочка заострённая. Уши крупные, без кисточек. Глаза большие. Мех короткий, сверху серого или коричневого цвета, животик белый, чёрная полоса проходит от глаза и чуть дальше уха, кончик пушистого хвоста украшен белой кисточкой.
Обитает в лиственных лесах и садах Западной Европы.
Магрибская соня (Eliomys munbyanus)
Встречается в Алжире, Ливии, Марокко, Тунисе и Западной Сахаре, во влажных лесах и полупустынях.
Копетдагская или мышевидная соня (Myomimus personatus)
Длина тела около 8 см. Хвост в длину 5-6 см. Спинка серого цвета с размытой темной полосой. Животик белый.
Обитает в Туркмении и Иране.
Болгарская мышевидная соня (Myomimus roachi)
Распространена в Болгарии, Греции и Турции.
Иранская мышевидная соня (Myomimus setzeri)
Вид является эндемиком Ирана.
Селевиния (Selevinia betpakdalaensis)
Длина тела около 10 см, хвост в длину до 7,5 см, вес около 30-35 г. Шерсть густая, пышная. Хвост густо покрыт короткими волосками. Спинка серого цвета, животик беловатый. Уши крупные, зверек способен сворачивать их в трубочку и разворачивать, как веер.
Редкий вид, является эндемиком Казахстана, где живет в пустынях.
Японская соня (Glirulus japonicas)
Эндемик Японии. Распространен в умеренных лесах. Способен быстро бегать вниз головой и повисать на ветках.
Орешниковая соня (Muscardinus avellanarius)
Длина тела около 15 см, вес от 15 до 25 г. Хвост в длину от 6 до 8 см, с кисточкой. Мордочка притуплённая, уши маленькие, округлой формы, усы длинные. Спинка охристо-рыжего цвета, редко с красноватым оттенком, брюшко светлое с палевым оттенком. Горло, грудь и животик светлые. Пальцы белого цвета. Кончик хвоста тёмный или светлый.
Горло, грудь и животик светлые. Пальцы белого цвета. Кончик хвоста тёмный или светлый.
Встречается в широколиственных и смешанных лесах Европы и северной Турции.
Соня-полчок (Glis glis)
Самая крупная из сонь с длиной тела от 13 до 18 см, весом 150-180 г. Похожа на серую белочку, но на округлых ушах нет кисточек. Хвост в длину от 11 до 16 см, пушистый, сверху серого цвета, снизу белёсый. Мех пышный и пушистый, дымчато-серый или серовато-бурый на спинке и светло-серый, желтоватый или белый на животике. Окологлазные кольца темные, тонкие.
Ареал обитания включает равнинные и горные леса Европы, Кавказа и Закавказья. Встречается на островах Средиземного моря: Сардинии, Корсике, Сицилии, Крите и Корфу.
Самец и самка сони: основные отличия
Половой диморфизм у сони не выражен. Самцы и самочки выглядят одинаково.
Поведение сони
Большинство сонь являются лесными животными и ведут ночной образ жизни. Также живут в лесостепи, в горах на высоте до 3500 м над уровнем моря, пустынях, саваннах. Некоторые сони (соня-полчок) практически все свое время проводят на деревьях, другие (мышевидные сони) живут на земле. Убежища сони обустраивают в дуплах, гнёздах, в норках, под корнями деревьев.
Размножение сони
Свои гнёзда круглой формы из листвы и травы сони располагают в кроне деревьев на высоте около 3 м над землёй. Их активно защищают самки. За год самки приносят один-два помёта. Продолжительность беременности 22-28 дней. В одном выводке от 2 до 9 малышей. Самостоятельными они становятся спустя месяц.
Продолжительность жизни сони в природе от 2 до 5,5 лет.
Естественные враги
На сонь охотятся ласки, горностаи, лесные куницы и другие хищные животные. Большой вред грызунам наносят паразиты, например, блохи.
Интересные факты о грызуне:
- Ближе к осени сони отъедаются и на зиму впадают в спячку, которая продолжается до полугода.
 Именно эта привычка послужила причиной названия вида.
Именно эта привычка послужила причиной названия вида. - Если человек или хищник ловят соню и случайно хватают зверька за хвост, то шкурка на нём разрывается и слезает словно чулок, а соня спасается бегством. Затем оголенный кончик хвоста ссыхается и мертвая часть отпадает. Со временем конец хвоста расширяется и обрастает шерстью.
- Иногда сони наносят ущерб садовым культурам.
- Шкурки крупных видов сонь заготавливаются как второстепенная пушнина.
 Именно эта привычка послужила причиной названия вида.
Именно эта привычка послужила причиной названия вида.ADW: Dryomys nitedula: ИНФОРМАЦИЯ
Географический ареал
Сонь лесная (Dryomys nitedula) встречается в Палеарктике. Они распространены по всей Европе и распространяются на юг до Северной Африки и на восток до Японии. (Haberl, 1999)
Среда обитания
Сонь лесная встречается в густых лесах, обычно в лиственных и смешанных лесах, а также в зарослях на высотах до 3500 м.Лесные сони используют возделываемые территории, такие как сады, а также каменистые луга. Для гнезда они выбирают густой кустарник или нижние ветви деревьев. (Хаберл, 1999; Кашталян, 1999)
- Диапазон превышения
- 3500 (выс.) М
- футов
Физическое описание
Длина головы и тела лесного сони от 80 до 130 мм.Длина хвоста колеблется от 60 до 113 мм. Масса тела колеблется от 18 до 34 г. Считается, что Дриомис очень похож на Элиомиса, но меньше по размеру. Мозг лесного сони более округлый, а слуховые буллы меньше по сравнению с Eliomys. (Haberl, 1999)
Лесные сони по внешнему виду напоминают белку, с серовато-коричневой или желтовато-коричневой спинкой и желтовато-белой нижней стороной. У них плоский и пушистый хвост, более однородный по цвету, чем у Элиомиса. (Haberl, 1999)
У них плоский и пушистый хвост, более однородный по цвету, чем у Элиомиса. (Haberl, 1999)
- Масса диапазона
- от 18 до 34 г
- 0.От 63 до 1,20 унции
- Длина диапазона
- от 80 до 130 мм
- от 3,15 до 5,12 дюйма
Репродукция
Информации о системе спаривания этого вида немного.
Сезон размножения лесной сони варьируется в зависимости от ареала вида.В Израиле сезон размножения длится с марта по декабрь. В среднем каждая самка рожает 2–3 раза в год. В Европе сезон размножения длится с мая по август, и обычно бывает только один помет в год. Период беременности составляет от 21 до 30 дней. Обычно в помете рождается от 2 до 5 особей, хотя иногда возможно и до 7 особей. (Haberl, 1999)
Каждое потомство при рождении весит примерно 2 г. Глаза не открываются примерно до 16-го дня жизни, а независимость от матери достигается только в возрасте от 4 до 5 недель.В Европе молодые будут ждать, пока не наступит первая зима, чтобы спариться. (Haberl, 1999; Nowak, 1999)
- Период размножения
- В Израиле вид будет размножаться 2–3 раза в год. В Европе вид размножается только один раз в год.
- Сезон размножения
- В Израиле брачный период длится с марта по декабрь.В Европе брачный период происходит с мая по август.
- Диапазон количества потомков
- от 2 до 7
- Среднее количество потомков
- 3
- Возраст
- Диапазон срока беременности
- от 21 до 30 дней
- Диапазон времени до независимости
- от 4 до 5 недель
- Средний возраст половой или репродуктивной зрелости (женщины)
- 1 год
- Средний возраст половой или репродуктивной зрелости (мужчины)
- 1 год
Информации о родительской заботе об этом виде мало. Новорожденные альтрицы и не открывают глаза, пока им не исполнится 16 дней. Молодые люди находятся на попечении матери до 4–5-недельного возраста. Пока они не станут независимыми, мать, скорее всего, будет обеспечивать их пищей (молоком), уходом и защитой. О родительской заботе со стороны мужчин не сообщается, но нельзя исключать и этого. (Haberl, 1999; Nowak, 1999)
Новорожденные альтрицы и не открывают глаза, пока им не исполнится 16 дней. Молодые люди находятся на попечении матери до 4–5-недельного возраста. Пока они не станут независимыми, мать, скорее всего, будет обеспечивать их пищей (молоком), уходом и защитой. О родительской заботе со стороны мужчин не сообщается, но нельзя исключать и этого. (Haberl, 1999; Nowak, 1999)
- отсутствие родительского участия
- альтриальный
- предварительное оплодотворение
- до вылупления / рождения
- перед отъемом / оперением
- до обретения независимости
Срок службы / Долговечность
Ожидается, чтоDryomys nitedula доживет до 5 особей.5 лет в дикой природе. (Haberl, 1999)
Поведение
Лесные сони очень древесные. Они обладают способностью подниматься с большой ловкостью, а также могут прыгать с ветки на ветку на высоту до 2 метров. (Haberl, 1999)
Этот вид строит на деревьях. Эти гнезда обычно собираются группами. Лесные сони обычно собирают временные гнезда, которые зачастую плохо построены и непрочны. Гораздо больше энергии вкладывается в строительство очень прочных родовых гнезд.Обычно они находятся на высоте 1–7 м над уровнем земли и имеют диаметр от 150 до 250 мм. Эти гнезда имеют сферическую форму с одним входом, обычно обращенным к стволу дерева. Гнезда строятся из листьев и веток и выстилаются фрагментами коры или мха. (Хаберл, 1999; Кашталян, 1999)
(Хаберл, 1999; Кашталян, 1999)
Dryomys nitedula ведет ночной образ жизни и проявляет гибернацию, а также дневную изнеможение. В Израиле эти животные остаются активными круглый год даже на больших высотах. Они действительно впадают в оцепенение зимой на определенное время каждый день.В северных частях своего распространения, например, в Европе, лесной сони впадают в спячку с октября по апрель. Во время спячки соня будет сидеть на задних лапах, свернуться клубком, обхватить тело хвостом и прижать руки к щекам. Было замечено, что они иногда появляются, чтобы поесть из запасов пищи. В России считается, что они активны всю зиму. (Haberl, 1999)
Лесные сони очень территориальные, с размерами территории от 65 до 100 м в диаметре.2
Домашний диапазон
Эти животные обычно занимают территории диаметром от 65 до 100 м. (Мак, 2001; Новаковски, 1999)
Коммуникация и восприятие
Дриомис издает различные вокализации. Наиболее заметным из них является тонкий мелодичный писк, который действует как сигнал будильника. Исследования содержащихся в неволе особей показали, что D.nitedula обладает способностью издавать повторяющиеся серии ультразвуковых исследований. Сигналы подавались обоими полами в ситуациях, предполагающих социальный характер общения. Эти сообщения были совершенно не слышны человеческому уху. (Боратынский и др., 1999; Haberl, 1999)
Хотя это специально не сообщается для этого вида, вполне вероятно, что тактильные, химические и визуальные сигналы являются частью репетуара общения. Млекопитающие обычно используют тактильное общение во время спаривания, конфликтов и воспитания детенышей. Химическая коммуникация может быть важна для индивидуальной идентификации, а также в репродуктивном контексте. Визуальные сигналы часто подаются посредством позирования тела, чтобы указать на враждебные или дружеские намерения.
Химическая коммуникация может быть важна для индивидуальной идентификации, а также в репродуктивном контексте. Визуальные сигналы часто подаются посредством позирования тела, чтобы указать на враждебные или дружеские намерения.
Привычки питания
Лесные сони всеядны. Они едят листья, а также отборные цветы, фрукты и орехи. Они также едят членистоногих, яйца и молодых птиц. Считается, что в течение лета предпочтительным диетическим продуктом является животный материал.(Haberl, 1999; Mack, 2001)
- птицы
- яйца
- насекомые
- наземные членистоногие, не являющиеся насекомыми
- листья
- семена, зерна и орехи
- фрукты
Хищничество
Dryomys nitedula ведет ночной образ жизни, встречается при низкой плотности, и особи очень осторожны, чтобы не покидать заросший кустарником покров.Такое поведение делает этих животных трудной добычей. Остатки лесного сони были обнаружены в гранулах сов, таких как Strix aluco и Bubo bubo, в Восточной Европе. (Мак, 2001; Обуч, 1999)
Роли экосистемы
Лесные сони могут играть роль в контроле за размножением членистоногих, составляющих значительную часть их рациона. Они также едят семена и фрукты и поэтому могут способствовать распространению семян. Поскольку этот вид дает пищу хищникам, таким как совы, лесные сони могут иметь некоторое положительное влияние на популяции этих хищников.(Haberl, 1999; Mack, 2001)
Поскольку этот вид дает пищу хищникам, таким как совы, лесные сони могут иметь некоторое положительное влияние на популяции этих хищников.(Haberl, 1999; Mack, 2001)
Экономическое значение для людей: положительный результат
Нет конкретных данных о положительном экономическом значении D. nitedula для человека.
Экономическое значение для людей: отрицательно
Известно, что популяции D. nitedula наносят экономический ущерб, совершая набеги на фруктовые сады и грызущие кору хвойных деревьев. (Haberl, 1999)
Статус сохранения
Dryomys nitedula считается исчезающим видом в Чешской Республике и редким в большинстве других европейских стран.МСОП определил лесных соней как менее опасных, но этот вид не внесен в список СИТЕС. Угроза для популяции в основном связана с разрушением лесной среды обитания на всей территории их ареала. (Haberl, 1999)
Великобритания разработала Национальную программу мониторинга сони, которая существует в первую очередь для мониторинга лещинных соней. Орешник - любимая звезда детских сборников сказок в Англии и Уэльсе, но элементы программы являются стандартными для всех видов сони. Исследователи поместили ящики для гнезд для сони на деревьях в лесных районах, где, как известно, встречаются сони, и возвращались в эти ящики, чтобы посчитать, определить пол и взвесить сони.Исследования проводятся с целью формирования базы данных по видам и мониторинга здоровья популяции. (Haberl, 1999)
English Nature проводит программу реинтродукции, в рамках которой с 1994 года повторно вводятся выращиваемые в неволе сони в районы, где когда-то было много популяций. Кроме того, английское правительство начало поощрять фермеров пересаживать живые изгороди, которые очень важны для среды обитания лесных соней. Такие усилия по сохранению также будут полезны для сохранения популяций других видов сони, таких как D.nitedula. (Мак, 2001)
Такие усилия по сохранению также будут полезны для сохранения популяций других видов сони, таких как D.nitedula. (Мак, 2001)
Недавно была обнаружена окаменелость сонеподобного млекопитающего, который считается самым ранним предком эвтериана. Окаменелость была полностью найдена в очень хорошо сохранившемся дне озера в Китае. Eomaia, название, данное окаменелости, означающее «древняя мать», обладает скелетными чертами, более близкими к современным плацентарям, чем к сумчатым. Это означает, что раскол между двумя группами произошел более 125 миллионов лет назад, до появления Эомайи.До того, как была обнаружена Эомаиа, самая старая зарегистрированная окаменелость плацентарного млекопитающего была зубами возрастом 110 миллионов лет, а самому старому черепу и скелету было всего 75 миллионов лет. (Hecht, 2002)
Авторы
Нэнси Шефферли (редактор), Animal Diversity Web.
Кимберли Скахан (автор), Университет Висконсина-Стивенс-Пойнт, Крис Янке (редактор, преподаватель), Университет Висконсина-Стивенс-Пойнт.
Глоссарий
- Палеарктика
проживает в северной части Старого Света.Другими словами, Европа и Азия и Северная Африка.
- акустический
использует звук для связи
- сельское хозяйство
живут на ландшафтах, где преобладает сельское хозяйство.
- альтернативный
человек родились в относительно слаборазвитом состоянии; они не могут кормиться, заботиться о себе или самостоятельно передвигаться в течение определенного периода времени после рождения / вылупления.
У птиц голые и беспомощные после вылупления.
 У птиц голые и беспомощные после вылупления.
У птиц голые и беспомощные после вылупления.- древесный
Обозначает животное, живущее на деревьях; лазание по деревьям.
- двусторонняя симметрия
, имеющий такую симметрию тела, что животное можно разделить в одной плоскости на две зеркальные половины. У животных с двусторонней симметрией есть спинная и вентральная стороны, а также передний и задний концы. Синапоморфия билатериев.
- химическая
использует запахи или другие химические вещества для общения
- эндотермический
животных, которые используют выделяемое метаболическим путем тепло для регулирования температуры тела независимо от температуры окружающей среды.Эндотермия - это синапоморфия млекопитающих, хотя она могла возникнуть у (ныне вымершего) предка синапсидов; летопись окаменелостей не различает эти возможности. Сходится у птиц.
- удобрения
союз яйцеклетки и сперматозоида
- лес
- В
лесных биомах преобладают деревья, в противном случае лесные биомы могут сильно различаться по количеству осадков и сезонности.

- гибернация
, состояние, в котором некоторые животные входят зимой, когда нормальные физиологические процессы значительно замедляются, что снижает потребность животных в энергии. Акт или условие прохождения зимы в оцепенении или состоянии покоя, обычно предполагающее отказ от гомойотермии у млекопитающих.
- итеропарная
потомков производятся более чем в одной группе (пометы, клатчи и т. Д.).) и в течение нескольких сезонов (или других периодов, благоприятных для размножения). По определению, итеропородящие животные должны выживать в течение нескольких сезонов (или периодических изменений состояния).
- подвижный
, имеющий возможность перемещаться с одного места на другое.
- родной диапазон
район, в котором животное обитает в естественных условиях, регион, в котором оно является эндемиком.
- ночной образ жизни
активен ночью
- всеядность
животное, которое в основном ест всевозможные продукты, включая растения и животных
- Сезонное разведение
разведение приурочено к определенному сезону
- малоподвижный
остается на том же участке
- половой
воспроизводство, включающее сочетание генетического вклада двух особей, мужчины и женщины
- тактильные
использует прикосновение для связи
- умеренный
этот регион Земли между 23.
5 градусов северной широты и 60 градусов северной широты (между тропиком Рака и Северным полярным кругом) и между 23,5 градусами южной широты и 60 градусами южной широты (между тропиком Козерога и Северным полярным кругом).
 5 градусов северной широты и 60 градусов северной широты (между тропиком Рака и Северным полярным кругом) и между 23,5 градусами южной широты и 60 градусами южной широты (между тропиком Козерога и Северным полярным кругом).
5 градусов северной широты и 60 градусов северной широты (между тропиком Рака и Северным полярным кругом) и между 23,5 градусами южной широты и 60 градусами южной широты (между тропиком Козерога и Северным полярным кругом).- наземный
Живут на земле.
- территориальный
защищает территорию в пределах ареала обитания, занятую отдельными животными или группой животных одного вида и удерживаемую посредством открытой защиты, демонстрации или рекламы
- УЗИ
использует звук, превышающий диапазон человеческого слуха, либо для навигации, либо для связи, либо для того и другого.
- визуальный
использует зрение для связи
- живородящие
размножение, при котором оплодотворение и развитие происходят в женском теле, а развивающийся эмбрион получает питание от самки.
Список литературы
П. Боратынский, А. Рахвальд, В. Новаковски. 1999. Ультразвуковые коммуникативные вызовы у лесной сони (* Dryomys nitedula *). IV Международная конференция по сони (Rodentia, Gliridae).
Доступ
01.11.02
на http://www.trakya.edu.tr/conference/d13. htm.
htm.
Haberl, W. 1999. "Сонная лощина: Дремыс" (В сети ).Сонная лощина. Доступ 20.10.2002 на http://www.gliratium.org/dormouse.
Hecht, J. 2002. Family Treat. New Scientist, 174/2340: 14.
Кашталян А. 1999. Сонь Беларуси: экология, распространение и история изучения. IV Международная конференция по сони (Rodentia, Gliridae). Доступ 01 ноября 2002 г. на http://www.trakya.edu.tr/conference/df6.htm.
Мак, Т. 2001. Восстание сони. Международная дикая природа, 31: 38.
Новак Р. 1999. Млекопитающие Уокера в мире, шестое издание. Балтимор и Лондон: Издательство Университета Джона Хопкинса.
Новаковски В. 1999. Система пространственного распространения * Dryomys nitedula * в Беловежской пуще (Восточная Польша). IV Международная конференция по сони (Rodentia, Gliridae). Доступ 01.11.02 по адресу http: // www.trakya.edu.tr/conference/d14.htm.
Обуч, Дж. 1999. Сони в рационе сов на Ближнем Востоке. IV Международная конференция по сони (Rodentia, Gliridae). Доступ 01.11.02 на http://www.trakya.edu.tr/conference/d21.htm.
Соня лесная (Dryomys nitedula) - life4oakforests.eu
Описание видов и местообитаний
Длина тела 15-24 см, из которых длина хвоста 6-11 см.Цвет его красновато-коричневый. Этот вид носит черную маску от носа до ушей. Лесная соня поедает животных, особенно насекомых. Встречается в горных лесах. Его среда обитания представляет собой богатые видами лиственные леса, особенно в ксеротермных, дубовых лесах, где кустарниковый ярус и опушки леса хорошо развиты, но лес не закрыт. Также его можно найти в соседних заброшенных садах.
Основные угрозы
Основная угроза для лесной сони - вырубка смешанных видов, исчезновение диких плодов и уничтожение кустарников и опушек кустарниковых лесов. Кроме того, сбор мертвых деревьев и кустарников сокращает среду обитания, а также создает однородные дубы. Кроме того, вырубка кустарниковых местообитаний, вырубка разнообразных дубовых и лиственных лесов, а также создание больших однородных лесов и вырубок снижают выживаемость этого вида.
Кроме того, сбор мертвых деревьев и кустарников сокращает среду обитания, а также создает однородные дубы. Кроме того, вырубка кустарниковых местообитаний, вырубка разнообразных дубовых и лиственных лесов, а также создание больших однородных лесов и вырубок снижают выживаемость этого вида.
Специальные консервационные мероприятия
Мы можем обеспечить подходящие повадки для этого вида, увеличив видовой состав опушек и ярусов кустарников.Также, открыв закрытый полог дубового леса, создав солнечные поляны разного размера, мы сможем облегчить появление цветущих кустарников. Улучшение видового состава древесных пород может быть достигнуто за счет стимулирования развития диких плодовых деревьев и других редких видов, а также за счет посадки кустарников на полянах и опушках леса. Создание различных видов валежника и установка искусственных скворечников еще больше способствует сохранению этого вида.
Области связанных проектов
fóti-Somlyó
Библиография
1. Хеккер, К. (2013): Популярный вид с высоты птичьего полета (Gliridae), открытый для наблюдения за биологией в Немзети. (Doktori értekezés) - Szent István Egyetem Környezettudományi Doktori Iskola, Gödöllő, 111 стр.
2. http://tolgy.tycmo.hu/peleeh.php
(PDF) Филогеография лесной сони Dryomys nitedula (Gliridae, Rodentia) Русской равнины и Кавказа
363
вдвое меньше D.nitedula - E. quercinus расстояние
(20,4 ± 0,4%, значение p <0,001). Для большей надежности
мы рассчитали p-расстояния между группами,
, на основе данных по аминокислотам (dAA). Они находились в
соответствии с T3P-расстояниями: Восточноевропейские -
Расстояния гаплогрупп кавказцев составляли 25,4 ± 2,9%
и 26,6 ± 2,9%, вдвое меньше D. nitedula - E.
quercinus p-distance (dAA = 45,5 ± 1,0%).
Западно-кавказская (Даховская, Архыз, Утриш,
Армавир) и Центрально-Кавказская (Стрижамент,
Учкулан, Эльбрус) гаплогруппы
можно рассматривать как родственные. Генетическая дистанция между ними составляет
Генетическая дистанция между ними составляет
значимых, d = 6,0 ± 0,9% (p-value <0,001), dAA =
16,1 ± 2,4%.
Ближайшие точки сбора гаплогрупп Западной и
Центрального Кавказа находятся на расстоянии
60 км без признаков гибридизации по гену cytb.
Вероятно, поток генов между этими гаплогруппами составляет
, прерванный географической или репродуктивной изоляцией.
Обсуждение
Современная внутривидовая таксономия D.nitedula - это исключительно
на основе особенностей окраски. В последней редакции
географическая изменчивость и систематика D. nitedula,
сони Русской равнины и Западного Кавказа
, распространенные на восток примерно до 42 ° в.д., относятся к номинативному подвиду
(Россолимо, 1971).
Предположительно на этой долготе на Кавказе граница
номинативного подвида и Dryomys nitedula
ognevi Heptner & Formosov, 1928 (подвид из
Южного Дагестана, син.caucasicus) проходит.
Предполагаемая граница Центральной и Западной
Кавказских гаплогрупп совпадает с одной из
номинативных подвидов и D. n. Огневи и с ландшафтами
Западного и Восточного Кавказа (Соколов
и Темботов 1989).
Наблюдаемый уровень генетической дивергенции между
русской равнинной и кавказской популяциями D. nitedula
достаточно высок. Это позволяет предположить, что имелось
длительной изоляции европейского и кавказского
ареалов D.nitedula на протяжении всего плейстоцена.
Три обнаруженные гаплогруппы D. nitedula отличаются
высокими уровнями нуклеотидов и аминокислот гена цитохрома
b. Родственные виды млекопитающих, определенные по морфологическим различиям
, обычно имеют более 5%
значений генетических дистанций cytb, а генетическая дистанция
, близкая к 10%, типична для «хороших» видов (Bradley &
Baker 2001). В любом случае, с подтверждением статуса вида следует быть осторожнее
. Поэтому очень важно исследовать
Поэтому очень важно исследовать
других популяций и
других генетических маркеров, чтобы подтвердить их особый статус как
, это было выполнено Григорьевой и соавт. (2015).
Благодарности
Работа поддержана грантами РФФИ 12-04-32035-мол_а,
12-04-00551 и грантом Президента РФ МК-3755.2014.4.
Таблица 1. Количество особей (N), географическое происхождение, выборка и названия гаплотипов D. nitedula.
Номер на карте и место происхождения N Номер ваучера. Гаплотипы GenBank
№ оценки
1 Белоруссия, Красное 51 ° 30′40 ′ ′ с.ш., 30 ° 30′10 ′ ′ в.д. 2 Бел114, Бел115 БЕЛ KJ739693
2 Россия, Саратов 51 ° 41′00 ′ ′ с.ш., 44 ° 53′51 ′ ′ E 2C5
C6
SR5
SR6
KF699220
KF699221
3 Россия, Саратов 51 ° 17′37 ′ ′ N, 44 ° 59′40 ′ ′ E 1
3
3
C , C69, C80, C83
C67-68, C78-79, C81-82
SR61
SR67
KF699223
KF699226
4 Россия, Саранск 54 ° 10′30 ′ ′ с.ш., 46 ° 9 ′ 59 ′ ′ E 5 S1, S2, S4, S5, S7 SN KJ739694
5 Россия, Серашмович 49 ° 33′26 ′ ′ N, 42 ° 39′48 ′ ′ E 7LD24, LD33
LD25-LD27, LD37, LD39
V24
V25
KJ739695
KJ739696
6 Россия, Серазмович 49 ° 42′15 ′ ′ с.ш., 42 ° 47′52 ′ ′ E 7
RD58, RD60, RD62, RD65 , RD66
RD57
RD59
V25
V57
V59
KJ739696
KJ739697
KJ739698
7 Россия, Утриш 44 ° 42′19 ″ с.ш., 37 ° 28′16 ″ в.д. 3 UTR1-3 UTR KJ739699
8 Россия, Даховская 44 ° 10′58 ″ с.ш., 40 ° 08′48 ″ в.д. 2ДАх2
ДАх3
ДАх2
ДАх3
KJ739700
KJ739701
9 Россия, Армавир 44 ° 57′21 ″ с.ш., 41 ° 10′07 ″ в.д. 2 ARM1-2 ARM KJ739708 ARM1-2 ARM KJ739708 ° 48′33 ″ с.ш., 42 ° 02′01 ″ в.д. 1 STR STR KJ739703
11 Россия, Эльбрус 43 ° 33′17 ″ с.ш., 42 ° 07′56 ″ в.д. 1ELBR ELBR KJ739704
12 Россия, г. Учкулан 43 ° 27′15 ″ с. ш., 42 ° 05′26 ″ в.д. 6 УЧ2-6 УЧ KJ739705
ш., 42 ° 05′26 ″ в.д. 6 УЧ2-6 УЧ KJ739705
13 Россия, Архыз 43 ° 33′57 ′ ′ с.ш., 41 ° 16′44 ′ ′ в.д. 1 0-22 0-22 AJ225116
(PDF) Экология лесной сони Dryomys nitedula (Pallas 1778) на северо-западном краю ареала
8 R.Юш кайтис: Экология лесной сони на краю ареала
Литература
Айрапетянц, А.Э. 1983. Соня. Издательство Ленинградского
университет, Ленинград. С. 191.
Angermann, R. 1963. Zur Ö kologie und Biologie des Baumschläfers,
Dryomys nitedula (Pallas, 1779) in der Waldsteppenzone. Acta
Териол. 7 (18): 333 - 367.
Бацайхан Н., Б. Крыш туфек, Г. Амори и Н.Йигит. 2008. Dryomys
nitedula. МСОП 2013. Красный список МСОП видов, находящихся под угрозой исчезновения.
Версия 2013.2. Доступно на: www.iucnredlist.org.
Давыдов Г.С. 1984. Распространение и экология леса.
Соня (Dryomys nitedula Pallas, 1779) в Таджикистане.
Известия Академии Наук Таджикской ССР, Биол. 2 (95): 55-60
(на русском языке).
Дума, I. 2013. Блоховая нагрузка и ее влияние на выбор гнезда
и использование в Dryomys nitedula Pallas, 1778.Acta Theriol. 58:
419–423.
Дума, И. и С. Джурджу. 2012. Циркадная активность и гнездо используют
Dryomys nitedula по данным инфракрасного датчика движения
камер. Folia Zool. 61: 49 - 53.
Флауэрдью Дж. Р. 1976. Экологические методы. Млекопитающее Rev.6: 123 - 159.
Голодшко, Б.З. и Э.Е.Падутов. 1961. Материалы по экологии
лесной сони Беловежской пущи. В: (N.D.
Ges ’, ed.) Фауна и экология наземных позвоночных
Белоруссии. Издательство Министерства высшего, среднего и
профессионального образования БССР, г. Минск. С. 49 - 70 (на
рус.).
Гански И. и М. Гилпин. 1991. Динамика метапопуляции: краткая история
и концептуальная область. Биол. J. Linn. Soc. 42: 3 - 16.
Homolka, M. 1979. Zur Alterbestimmung der Schläfer (Gliridae).
Folia Zool. 28: 103 - 114.
Юшкайтис, Р. 2005. Влияние высокой плотности гнездовых яиц на популяцию обыкновенной сони Muscardinus avellanarius (
). Acta
Териол. 50: 43 - 50.
Юшкайтис Р. 2008. Соня обыкновенная Muscardinus
avellanarius: экология, структура и динамика популяции.
Институт экологии издательства Вильнюсского университета, Вильнюс.
стр. 163.
Юшкайтис Р. 2010. Обитатели гнездовий: птицы, млекопитающие, социальные
насекомых.Лютут Э, Каунас. С. 160 (на литовском языке с резюме на английском языке
).
Юш Кайтис, Р. и Л. Балтрунаит Э. 2013. Сезонная изменчивость рациона лесной сони Dryomys nitedula
на северо-западном краю ареала распространения
. Folia Zool. 62:
311 - 318.
Юш Кайтис, Р., Л. Балчяускас и В. Шио ж иныт э. 2012. Место гнездования
предпочтение лесной сони Dryomys nitedula (Pallas) в северо-западном углу ареала
.Pol. J. Ecol. 60:
815 - 826.
Кребс, Ч. Дж. 1999. Экологическая методология, 2-е изд. Эддисон Уэсли
Лонгман, Менло-Парк, Калифорния. С. 620.
Крыш туфек, Б. 1985. Соня лесная Dryomys nitedula (Pallas,
1778) - Rodentia, Mammalia - в Югославии. Scopolia.
9: №1 - 36.
Крыш туфек, Б. 2010. Glis glis (Rodentia: Gliridae). Мамм. Виды
42: 195 - 206.
Kry š tufek, B.и В. Вохралик. 1994. Распространение лесной сони
, Dryomys nitedula (Pallas, 1779) (Rodentia:
Myoxidae) в Европе. Mammal Rev.24: 161 - 177.
Lozan, M.N. 1961. Определение возраста Dryomys nitedula Pall.
и Muscardinus avellanarius L. Zool. Ж. 40: 1740 - 1743
(на русском языке с резюме на английском языке).
Лозан М.Н. 1970. Грызуны Молдавии. Vol. 1. Редакционно-
издательский отдел Академии наук Молдавской ССР, Кишинев.
с. №168.
Лозан М., Белик Л., Самарский С. 1990. Сонь (Gliridae)
Юго-Запада СССР. Штиинца, Кишинев. С. 146.
Штиинца, Кишинев. С. 146.
Марков Г., Атанасова И., Райков И., Господинова М. 2009.
Демографическая структура населения лесной сони
(Dryomys nitedula Pall., 1779) в искусственной лесозащитной полосе
в Болгарии. Компт. Ренд. Акад. Bulg. Sci. 62: 485 - 490.
Maškov á, P. and P. Adam í k. 2012. Заметки о встречаемости
древесных грызунов (Mammalia: Rodentia) в гнездовьях в районе
Совинец, N í zk ý Jesen í k Mts., Чешская Республика. Zpr á vy
Vlastiv ě dn é ho muzea v Olomouci 303: 13 - 21 (на чешском языке с резюме на английском языке
).
Моррис, П.А. 2003. Обзор исследований британских соней (Gliridae)
и эффекта повышения общественной и научной осведомленности о
этих животных.Acta Zool. Подвешенный. 49 (Приложение 1): 125 - 130.
Morris, P.A., P.W. Брайт и Д. Вудс. 1990. Использование скворечников
соней Muscardinus avellanarius. Биол. Консерв. 51: 1–13.
Нево, Э. и Э. Амир. 1964. Географическая изменчивость репродукции
и режимы гибернации лесной сони. J. Mammal.
45: 69 - 87.
Новаковски, В.К. и П. Боратынски. 2001. Попытка оценить
размер и плотность популяции Dryomys nitedula в лесу
Bia łowie ż.Trakya Univ. J. Sci. Res., Ser. B. 2: 121 - 124.
Nowakowski, W.K. и М. Годлевска. 2006. Значение
корма для животных для Dryomys nitedula Pallas и Glis glis L. в
Bia łowie ż a Forest (Восточная Польша): анализ фекалий. Pol. J. Ecol.
54: 359 - 367.
Огнев С.И. 1947. Млекопитающие СССР и сопредельных стран.
Грызуны. Vol. 5. Издательство Академии наук СССР, Москва-
Ленинград. стр.809 с.
Panchetti, F., G. Amori, G.M. Карпането и А. Сораче. 2004. Деятельность
образцов сони обыкновенной (Muscardinus avellanarius)
в различных средиземноморских экосистемах. J. Zool. 262: 289 - 294.
Паолуччи П., А. Баттисти и Р. де Баттисти. 1987. Соня лесная
1987. Соня лесная
(Dryomys nitedula Pallas, 1779) в Восточных Альпах (Rodentia,
Gliridae). Биогеография 13: 855 - 866.
Пилатс, В., Д. Пилате, А. Орниканс и А. Карклинш. 2012. Использование Microhabitat
лесным сони (Dryomys nitedula) в борео-
неморальном лесу - предварительные результаты. Peckiana 8: 77 - 85.
Rossolimo, O.L. 1971. Изменчивость и таксономия Dryomys nitedula
Pallas. Zool. Ж. 50: 247 - 258 (на русском языке с аннотацией на английском языке
).
Россолимо, О.Л., Э.Г. Потапова, И.Я. Павлинов, С. Крускопа и
О.В.Вольцит. 2001. Сонь (Myoxidae) мира. Издательство
Московского университета, Москва. С. 229.
Самарский, С.Л. и, как. Самарский. 1979. Разведение лесной сони
в условиях среднего Днепра. Экология
(Свердловск) 3: 96 - 99.
Schedl, W. 1968. Der Tiroler Baumschläfer (Dryomys nitedula
intermediateus [Nehring, 1902]) (Rodentia, Muscardinidae). Ein
Beitrag zur Kenntnis seiner Verbreitung und Ö kologie.Бер.
нат-мед. Вер. Инсбрук. 56: 389 - 406.
Подтверждено | [email protected] авторская копия
Дата загрузки | 19.03.14, 6:26
Млекопитающие
Распространение карликовой летучей мыши Molossops temminckii (Chiroptera: Molossidae) в Колумбии и комментарии к ее морфометрии
Хулио Дж.Чакон-Пачеко, Луис Хосе Авенданьо Мальдонадо, Карлос Агамес-Лопес, Ингрит Юлиани Мехиа-Фонтеча, Даниэла Веласкес-Гуарин, Паула А. Осса Лопес, Фреди А. Ривера Паес, Дарвин М. Чавес-Мартин Э. 21 октября 2020 г.
Аннотация
Род Molossops включает два вида, обитающих в Южной Америке: Molossops neglectus и Molossops temminckii. Меньший карлик с собачьей мордой Molossops temminckii распространен от Колумбии до Аргентины и имеет широкие морфологические вариации и вокальную пластичность. В Колумбии этот вид остается малоизвестным. Чтобы заполнить пробелы в распределении, мы представляем новые записи из департаментов Араука, Атлантико, Боливар, Кордова и Уила. Мы также представляем анализ морфометрической изменчивости в Южной Америке с использованием анализа главных компонентов. Они показывают внешнее и черепное отличие особей Колумбии от других популяций и популяций Южной Америки.В колумбийских ландшафтах, где преобладает пойменная саванна региона Ориноко, Molossops temminckii меньше, чем в других регионах страны, и следует оценить ранее предполагаемое существование загадочного разнообразия внутри таксона. Поэтому мы предлагаем дальнейший интегративный анализ для изучения возможного подвидового статуса некоторых колумбийских популяций.
Меньший карлик с собачьей мордой Molossops temminckii распространен от Колумбии до Аргентины и имеет широкие морфологические вариации и вокальную пластичность. В Колумбии этот вид остается малоизвестным. Чтобы заполнить пробелы в распределении, мы представляем новые записи из департаментов Араука, Атлантико, Боливар, Кордова и Уила. Мы также представляем анализ морфометрической изменчивости в Южной Америке с использованием анализа главных компонентов. Они показывают внешнее и черепное отличие особей Колумбии от других популяций и популяций Южной Америки.В колумбийских ландшафтах, где преобладает пойменная саванна региона Ориноко, Molossops temminckii меньше, чем в других регионах страны, и следует оценить ранее предполагаемое существование загадочного разнообразия внутри таксона. Поэтому мы предлагаем дальнейший интегративный анализ для изучения возможного подвидового статуса некоторых колумбийских популяций.
популяций лесной сони (Dryomys nitedula) на юге Италии принадлежат к глубоко дивергентной эволюционной линии
Abstract
Лесная соня ( Dryomys nitedula ) - небольшой грызун с широким, хотя и сильно фрагментированным распространением, от центральной Европы до Центральная Азия.В итальянском регионе популяций D. nitedula ограничены лесными горными районами двух в значительной степени разобщенных регионов, восточных Альп и региона Калабрия, где находятся два разных подвида ( D. nitedula intermediateus и D. nitedula aspromontis , соответственно) были описаны на основе фенотипических признаков (например, цвета меха). Здесь мы проанализировали образцов D. nitedula из обоих регионов, чтобы исследовать закономерности генетической дивергенции и филогенетических отношений между этими двумя популяциями.Генетическая изменчивость изучалась на уровне одного митохондрия (ген цитохрома b) и трех фрагментов ядерного гена (экзон 1 интерстициального ретиноид-связывающего белка, экзон 10 рецептора гормона роста и ген 1, активирующий рекомбинацию). Филогенетический анализ проводился с использованием методов максимального правдоподобия и байесовского вывода. D. n. aspromontis и D. n. Intermedius оказались реципрокно монофилетическими во всех филогенетических анализах, и генетическая дивергенция, наблюдаемая между ними по митохондриальному гену CYTB , была заметной (HKY: 0.044) по сравнению с ранее наблюдаемыми значениями у многих сестринских видов грызунов. Наши результаты ясно показывают, что D. nitedula aspromontis является глубоко расходящейся, узкой эндемичной эволюционной линией, и в ближайшем будущем необходимо тщательно оценить потребности в ее сохранении. Более того, такое глубокое генетическое расхождение вместе с фенотипической дифференциацией между D. n. Intermedius и D. n. aspromontis, предполагают, что популяций D. nitedula на юге Италии могут принадлежать к отдельным, ранее нераспознанным видам.
Филогенетический анализ проводился с использованием методов максимального правдоподобия и байесовского вывода. D. n. aspromontis и D. n. Intermedius оказались реципрокно монофилетическими во всех филогенетических анализах, и генетическая дивергенция, наблюдаемая между ними по митохондриальному гену CYTB , была заметной (HKY: 0.044) по сравнению с ранее наблюдаемыми значениями у многих сестринских видов грызунов. Наши результаты ясно показывают, что D. nitedula aspromontis является глубоко расходящейся, узкой эндемичной эволюционной линией, и в ближайшем будущем необходимо тщательно оценить потребности в ее сохранении. Более того, такое глубокое генетическое расхождение вместе с фенотипической дифференциацией между D. n. Intermedius и D. n. aspromontis, предполагают, что популяций D. nitedula на юге Италии могут принадлежать к отдельным, ранее нераспознанным видам.
Введение
Итальянский полуостров долгое время считался одним из основных компонентов биоразнообразия в Западном Средиземноморье и важным ледниковым убежищем для видов животных умеренного пояса на протяжении плио-плейстоцена (Hewitt, 2011). Появление и широкое применение генетических маркеров для изучения географической изменчивости значительно улучшило наше понимание ключевых биогеографических закономерностей и исторических процессов в этой области, выявив динамику расширения-сокращения, фрагментацию населения на несколько плейстоценовых рефугиумов, скрытые гибридные зоны, а также появление множества загадочных и глубоко расходящихся эволюционных линий (Barbanera et al., 2009; Canestrelli et al., 2006a, 2006b, 2007a, 2007b, 2008a, 2010, 2012a, 2012b, 2014a, 2014b; Канестрелли и Насчетти, 2008; Castiglia et al., 2007, 2016; Коланджело и др., 2012; Гриль и др., 2009; Киндлер и др., 2013; Lecocq et al., 2013; Ло Брутто и др., 2010; Луи и др., 2013; Маура и др., 2014; Mezzasalma et al., 2015; Nascetti et al., 2005; Salvi et al. , 2013; Salvi et al., 2017; Симонсен и Хьюмер, 2014; Wauters et al., 2017).
, 2013; Salvi et al., 2017; Симонсен и Хьюмер, 2014; Wauters et al., 2017).
Лесная соня Dryomys nitedula (Pallas, 1778) - небольшой грызун с широким, хотя и фрагментированным географическим распространением, от восточной и южной Европы до Центральной Азии (Krystufek, Vohralik, 1994).Несмотря на широкое распространение, современные знания о его экологии и систематике скудны. Этот вид ведет древесный и ночной образ жизни, и его наблюдали от уровня моря до более 2000 м над уровнем моря в самых разных средах обитания, но с заметными различиями между местными популяциями (Krystufek and Vohralik, 1994; Paolucci et al. 1989; Амори и др., 2008). Вместе с широким, но географически структурированным разнообразием размеров тела, цвета шерсти и, в меньшей степени, морфологии, эти различия между местными популяциями привели нескольких авторов к предположению о возможном появлении скрытых видов в пределах D.nitedula (Холден, 2005). Хотя всестороннее исследование его молекулярной систематики все еще отсутствует, немногочисленные доступные данные, похоже, подтверждают эту гипотезу (например, Grigoryeva et al., 2015) и указывают на то, что внутри этого номинального вида могут существовать загадочные дивергентные линии.
На Итальянском полуострове популяций D. nitedula ограничены лесными горными районами двух в значительной степени разобщенных регионов: восточных Альп и южной Италии (горные массивы Аспромонте, Сила и Поллино).Однако в недавнем прошлом этот большой разрыв в распределении мог быть меньше. Данные по окаменелостям позволяют предположить, что этот вид обитал в центральной Италии, по крайней мере, до последней ледниковой фазы (65-35 тысяч лет назад; см. Kotsakis, 1991, 2003). Основываясь на различиях в окраске шерсти (Nehring, 1902; Von Lehmann, 1964), две популяции из восточных Альп и южной Италии до сих пор были описаны как два разных подвида D. nitedula intermediateus Nehring, 1902 и D.nitedula aspromontis Von Lehmann, 1964, соответственно, причем последний демонстрирует более яркий серый мех и отчетливое белое пятно на кончике хвоста (Von Lehmann, 1964). Несмотря на обширные фаунистические исследования в районе Калабрии (Aloise и Cagnin, неопубликованные данные), D. n. aspromontis особей до сих пор встречались только на высотах выше 1000 м над уровнем моря и только в лесах, где преобладали буковые ( Fagus sylvatica ) (Cagnin and Aloise, 1995), тогда как вдоль Альп этот вид также встречался на более низких высотах. , и в основном в смешанных лесах из широколиственных и хвойных пород (Паолуччи и др., 1989). Однако цитогенетических и морфометрических различий между обоими подвидами не наблюдалось (Civitelli et al., 1995; Filippucci et al., 1995), и сообщалось об ограниченной генетической дифференциации на основании предварительных данных по аллозимам (Filippucci et al., 1995) , что ведет к неуверенности в правильном таксономическом отнесении популяций на юге Италии (Amori et al., 2008).
Несмотря на обширные фаунистические исследования в районе Калабрии (Aloise и Cagnin, неопубликованные данные), D. n. aspromontis особей до сих пор встречались только на высотах выше 1000 м над уровнем моря и только в лесах, где преобладали буковые ( Fagus sylvatica ) (Cagnin and Aloise, 1995), тогда как вдоль Альп этот вид также встречался на более низких высотах. , и в основном в смешанных лесах из широколиственных и хвойных пород (Паолуччи и др., 1989). Однако цитогенетических и морфометрических различий между обоими подвидами не наблюдалось (Civitelli et al., 1995; Filippucci et al., 1995), и сообщалось об ограниченной генетической дифференциации на основании предварительных данных по аллозимам (Filippucci et al., 1995) , что ведет к неуверенности в правильном таксономическом отнесении популяций на юге Италии (Amori et al., 2008).
В этом исследовании мы исследуем закономерности генетической дивергенции между D. n. aspromontis и D.п. Intermedius путем анализа паттернов вариации последовательностей на уровне одного митохондриального и трех ядерных фрагментов гена. Наша цель состояла в том, чтобы лучше охарактеризовать филогенетические отношения между популяцией лесной сони на юге Италии и ее представителями вида на севере. Фактически, учитывая большой географический разрыв между подвидами, расселение и обмен генами выглядят довольно неправдоподобными. Следовательно, оценивая, соответствует ли Д. п. aspromontis можно точнее определить как ограниченно дифференцированный географический обособленный объект или как уникальное эволюционное происхождение, которое может иметь серьезные последствия не только для таксономии, но и как значительную природоохранную ценность.
Материалы и методы
Всего было проанализировано 15 образцов D. nitedula (см. Рисунок 1 и таблицу 1). Хвостовые образцы D. n. aspromontis (n = 8) были собраны в поле, поскольку этот вид использует аутотомию хвоста в качестве антихищного поведения (Mohr, 1941). В качестве альтернативы образцы тканей также были взяты у погибших на дорогах людей. Все образцы были доставлены в лабораторию и хранились в 95% этаноле до экстракции ДНК. Образцы тканей D.п. Intermedius (n = 7) были любезно предоставлены Научным музеем Тренто (Muse) и Университетом Падуи в качестве образцов, консервированных этанолом (см. Таблицу 1).
В качестве альтернативы образцы тканей также были взяты у погибших на дорогах людей. Все образцы были доставлены в лабораторию и хранились в 95% этаноле до экстракции ДНК. Образцы тканей D.п. Intermedius (n = 7) были любезно предоставлены Научным музеем Тренто (Muse) и Университетом Падуи в качестве образцов, консервированных этанолом (см. Таблицу 1).
Географическое положение 15 образцов Dryomysnitedula , проанализированных в данном исследовании.
Рис. 1.A) Географическое положение образцов Dryomys nitedula aspromontis , проанализированных для настоящего исследования; нумерация населенных пунктов указана в таблице 1. B) Географическое распространение Dryomys nitedula в Европе и соседних регионах (преобразовано из Juškaitis, 2014). C) Географическое положение образцов Dryomys nitedula intermediateus , проанализированных для настоящего исследования; населенные пункты пронумерованы, как в таблице 1. D-G) Филогенетические деревья, выведенные для четырех фрагментов генов, проанализированных на основе процедуры байесовского вывода; числа указывают байесовские апостериорные вероятности.
Рисунок 2.Изображения двух подвидов Dryomy snitedula , обитающих на итальянском полуострове. A) Dryomys nitedula aspromontis (Monte Altare Longobucco; Фото: A.Пеллегрино). B) Dryomy snitedula intermediateus (Фото: Л. Лапини).
Полную геномную ДНК экстрагировали с помощью универсального набора ZR (Zymo Research), следуя стандартному протоколу выделения ДНК. Частичные митохондриальные последовательности гена цитохрома b ( CYTB ) были получены с использованием следующих праймеров (Григорьева и др., 2015): F_Dr.n_cyt (TGACAAACATCCGTAAAACT) и R_Dr.n_int (AAAAGCGGGTTAGTGTTGC). Амплификации методом полимеразной цепной реакции (ПЦР) проводили с изменениями исходного протокола (Григорьева и др. , 2015): начальный этап денатурации при 94 ° C в течение 3 минут, затем 30 повторных циклов при 94 ° C в течение 30 секунд, 54 ° C в течение 30 секунд и 72 ° C в течение 1 минуты, а также один заключительный этап при 72 ° C в течение 5 минут. Были амплифицированы три фрагмента ядерного гена: экзон1 интерстициальный ретиноид-связывающий белок ( IRBP ), экзон 10 рецептора гормона роста ( GHR ) и часть гена 1 активации рекомбинации ( RAG1 ). Использованные праймеры для ПЦР и условия цикла были такими же, как представлены в Pisano et al., 2015. Амплификации проводили с использованием идентичных смесей ПЦР для всех проанализированных фрагментов генов, включая: 20 нг экстрагированной ДНК в 25-мкл реакционной смеси, содержащей MgCL2 (2,5 ммоль / л), реакционный буфер (1X; Promega), четыре dNTP (0,2 ммоль / л каждый), два праймера (0,2 мкмоль / л каждый) и фермент Taq-полимераза (1 единица; Promega). Продукты ПЦР очищали и секвенировали Macrogen Inc. (htpp: //macrogen.com) с использованием системы секвенирования ABI PRISM 3700.
, 2015): начальный этап денатурации при 94 ° C в течение 3 минут, затем 30 повторных циклов при 94 ° C в течение 30 секунд, 54 ° C в течение 30 секунд и 72 ° C в течение 1 минуты, а также один заключительный этап при 72 ° C в течение 5 минут. Были амплифицированы три фрагмента ядерного гена: экзон1 интерстициальный ретиноид-связывающий белок ( IRBP ), экзон 10 рецептора гормона роста ( GHR ) и часть гена 1 активации рекомбинации ( RAG1 ). Использованные праймеры для ПЦР и условия цикла были такими же, как представлены в Pisano et al., 2015. Амплификации проводили с использованием идентичных смесей ПЦР для всех проанализированных фрагментов генов, включая: 20 нг экстрагированной ДНК в 25-мкл реакционной смеси, содержащей MgCL2 (2,5 ммоль / л), реакционный буфер (1X; Promega), четыре dNTP (0,2 ммоль / л каждый), два праймера (0,2 мкмоль / л каждый) и фермент Taq-полимераза (1 единица; Promega). Продукты ПЦР очищали и секвенировали Macrogen Inc. (htpp: //macrogen.com) с использованием системы секвенирования ABI PRISM 3700.
Полученные последовательности проверяли визуально с помощью CHROMAS 2.31 (TechnelysiumLtd.), И они были согласованы с CLUSTALX (Thompson et al., 1997) с настройками по умолчанию. Все полученные последовательности были депонированы в базе данных GenBank (инвентарные номера: XXX-XXX [заполняется после принятия]). Разнообразие последовательностей и паттерны дивергенции между последовательностями оценивали с помощью DIVEIN (Deng et al., 2010). Ядерные гетерозиготные последовательности фазировали с использованием ФАЗЫ 2.1 (Stephens et al., 2003) с вариантами по умолчанию, тогда как возникновение рекомбинации оценивали с использованием индекса попарной гомоплазии (статистика PHI, Bruen et al., 2006) в SPLITSTREE v.4.11 (Huson and Bryant, 2006).
Наиболее подходящая модель эволюции последовательности была выбрана для каждого проанализированного фрагмента гена среди 88 альтернативных моделей с использованием байесовского информационного критерия (BIC) в JMODELTEST 2. 1.3 (Darriba et al., 2012). Этот метод предложил HKY как лучшую модель замены для митохондриального фрагмента ( CYTB ), HKY + I для гена IRBP и JC + I для генов GHR и RAG1 .
1.3 (Darriba et al., 2012). Этот метод предложил HKY как лучшую модель замены для митохондриального фрагмента ( CYTB ), HKY + I для гена IRBP и JC + I для генов GHR и RAG1 .
Филогенетические деревья оценивались с помощью алгоритма максимального правдоподобия (ML), реализованного в программе PhyML (Guidon et al., 2010), используя настройки по умолчанию для всех параметров, со следующими исключениями: i) поддержка узла оценивалась с помощью непараметрической процедуры начальной загрузки на основе 1000 псевдоповторных копий; ii) для каждого использовалась лучшая модель замещения, как указано JMODELTEST. анализируемый маркер. Чтобы проверить соответствие между различными процедурами оценки филогенетических деревьев, филогенетические деревья также оценивались на основе процедуры байесовского вывода (BI) с помощью программного обеспечения MRBAYES v.3.2.1 (Ronquist et al., 2012).С этой целью четыре цепи Монте-Карло Маркова были запущены для 10 миллионов поколений, при этом деревья отбирались каждые 1000 поколений, а первые 25% полученных деревьев отбрасывались как выгорающие.
Результаты
Для всех проанализированных индивидуумов были получены последовательности длиной 427 п.н. для фрагмента гена CYTB , 889 п.н. для GHR , 1216 для IRBP и 826 п.н. для RAG1. Митохондриальная область размером 427 п.н. CYTB показала 21 вариабельную позицию, 20 информативных экономичных, в то время как индели, стоп-кодоны и нонсенс-кодоны не наблюдались.Ген GHR показал 18 вариабельных положений, из которых 17 информативных положений, ген IRBP представил 29 вариабельных положений, из которых 24 информативных положения, а ген RAG1 показал 8 вариабельных положений, 6 из которых информативны. Тест PHI, проведенный с фрагментами ядерных генов, не дал статистически значимых указаний на события рекомбинации.
Поскольку филогенетические деревья, выведенные с помощью методов ML и BI, дали полностью совпадающие топологии деревьев, здесь будут представлены только результаты, основанные на BI (деревья ML доступны по запросу). Как показано на Рисунке 1, для всех проанализированных генетических маркеров топология дерева четко идентифицировала образцы, принадлежащие D. n. aspromontis (юг Италии) и D. n. Intermedius (северо-восток Италии) как два взаимно монофилетических и хорошо поддерживаемых клона, без экземпляров общего гаплотипа. Среднее расхождение последовательностей между гаплотипами внутри каждой группы было минимальным, и ниже значения наблюдались между группами по всем анализируемым маркерам (см. Таблицу 2). Наивысшее значение расхождения, оцененное между обеими группами (HKY = 0.044; p-distance = 0,043) наблюдали на фрагменте гена мтДНК ( CYTB ).
Как показано на Рисунке 1, для всех проанализированных генетических маркеров топология дерева четко идентифицировала образцы, принадлежащие D. n. aspromontis (юг Италии) и D. n. Intermedius (северо-восток Италии) как два взаимно монофилетических и хорошо поддерживаемых клона, без экземпляров общего гаплотипа. Среднее расхождение последовательностей между гаплотипами внутри каждой группы было минимальным, и ниже значения наблюдались между группами по всем анализируемым маркерам (см. Таблицу 2). Наивысшее значение расхождения, оцененное между обеими группами (HKY = 0.044; p-distance = 0,043) наблюдали на фрагменте гена мтДНК ( CYTB ).
Среднее расхождение последовательностей (оценка максимального правдоподобия) внутри и между основными группами гаплотипов, полученных с помощью филогенетического анализа, проведенного среди образцов D. nitedula , проанализированных в настоящем исследовании. Стандартные ошибки указаны в скобках.
Обсуждение
Исследования внутривидового разнообразия лесной сони почти полностью основывались на фенотипических моделях изменчивости (но см. E.грамм. Филиппуччи и др., 1995; Григорьева и др., 2015), и приводят к описанию нескольких подвидов в рамках номинального вида D. nitedula . Однако, насколько эти фенотипические варианты фактически являются эволюционно независимыми линиями, все еще остается в значительной степени неизвестным. В этом исследовании мы проанализировали закономерности генетической дивергенции между географически изолированными популяциями лесной сони на юге Италии ( D. n. Aspromontis ) и их географически ближайшей популяцией на северо-востоке Италии ( D.п. промежуточный ).
Наши результаты ясно показывают, что D. n. aspromontis - это независимая эволюционная единица, монофилетическая по всем анализируемым маркерам и сильно отличающаяся по мтДНК от географически ближайшей популяции в северо-восточной Италии (HKY = 0,044). Эти результаты, по-видимому, противоречат данным Filippucci et al. (1995), в которых предполагался довольно низкий уровень аллозимной дифференцировки (D = 0,03). Тем не менее, хотя некоторые расхождения с точки зрения генетического разнообразия и моделей дифференциации не вызывают удивления (Toews and Brelsford, 2012), прямое сравнение двух оценок расхождения вряд ли имеет смысл.В самом деле, учитывая полностью аллопатрическое распределение двух родословных, обсуждение возможного несоответствия могло быть основано только на сравнении показателей генетической дистанции, полученных из различных методологических подходов. Тем не менее, стоит отметить, что, хотя Филиппуччи и его коллеги (1995) не идентифицировали ни одного аллозимического локуса, имеющего полную диагностическую ценность между подвидами, наши результаты указали на идеальную реципрокную монофилию в трех изученных ядерных локусах, таким образом, предполагая отсутствие энергетического разрешения. аллозимических локусов, использованных Филиппуччи и его коллегами (1995).
Эти результаты, по-видимому, противоречат данным Filippucci et al. (1995), в которых предполагался довольно низкий уровень аллозимной дифференцировки (D = 0,03). Тем не менее, хотя некоторые расхождения с точки зрения генетического разнообразия и моделей дифференциации не вызывают удивления (Toews and Brelsford, 2012), прямое сравнение двух оценок расхождения вряд ли имеет смысл.В самом деле, учитывая полностью аллопатрическое распределение двух родословных, обсуждение возможного несоответствия могло быть основано только на сравнении показателей генетической дистанции, полученных из различных методологических подходов. Тем не менее, стоит отметить, что, хотя Филиппуччи и его коллеги (1995) не идентифицировали ни одного аллозимического локуса, имеющего полную диагностическую ценность между подвидами, наши результаты указали на идеальную реципрокную монофилию в трех изученных ядерных локусах, таким образом, предполагая отсутствие энергетического разрешения. аллозимических локусов, использованных Филиппуччи и его коллегами (1995).
Во время анализа данных мы воздержались от использования мтДНК для молекулярного датирования, поскольку неполный отбор таксонов может сильно повлиять на итоговые оценки (Poux et al., 2008; Nabhan and Sarkar, 2012), а также наши образцы из D. n . Intermedius был в основном неполным. Тем не менее, мы не можем не заметить, что дивергенция последовательностей, наблюдаемая на CYTB между северной и южной выборками, предполагает гораздо более раннюю дивергенцию для D. n. aspromontis , чем средний голоцен (приблизительно 10 000 лет) или поздний плейстоцен (35 000-65 000), как предполагалось ранее на основе морфологических и ископаемых данных, соответственно (Roesler and Witte, 1968; Krystufek and Vohralik, 1994; Filippucci et al., 1995). Фактически, используя недавно оцененную частоту мутаций 0,0217 мутаций / сайт / миллион лет для CYTB у млекопитающих (Igea et al., 2015), величина дивергенции последовательностей, которую мы обнаружили, позволяет предположить, что время дивергенции между подвидами составляет около 1 миллиона. лет назад (т. е. ранний плейстоцен), таким образом, предшествовал этому событию по сравнению с предыдущими оценками. Следовательно, единственная летопись окаменелостей D. nitedula , обнаруженная в центральной Италии (Kotsakis, 1991, 2003), может свидетельствовать о недавнем сокращении ареала обитания в южной Италии бывшей «полуостровной» линии, как уже было показано для большого количества животных. виды в районе, (e.грамм. Канестрелли и др., 2006а; 2008; Гриль и др., 2009; Castiglia et al., 2016; Colangelo et al., 2012), а не совсем недавнюю (т.е. от позднего плейстоцена до середины голоцена) колонизацию южной Италии из Альп, как считалось ранее (Roesler and Witte, 1968; Krystufek and Vohralik, 1994; Filippucci et al., 1995). ).
лет назад (т. е. ранний плейстоцен), таким образом, предшествовал этому событию по сравнению с предыдущими оценками. Следовательно, единственная летопись окаменелостей D. nitedula , обнаруженная в центральной Италии (Kotsakis, 1991, 2003), может свидетельствовать о недавнем сокращении ареала обитания в южной Италии бывшей «полуостровной» линии, как уже было показано для большого количества животных. виды в районе, (e.грамм. Канестрелли и др., 2006а; 2008; Гриль и др., 2009; Castiglia et al., 2016; Colangelo et al., 2012), а не совсем недавнюю (т.е. от позднего плейстоцена до середины голоцена) колонизацию южной Италии из Альп, как считалось ранее (Roesler and Witte, 1968; Krystufek and Vohralik, 1994; Filippucci et al., 1995). ).
Наши результаты имеют большое значение для сохранения лесных соней на юге Италии. Фактически, наши результаты определенно идентифицируют эту линию как уникальную эволюционно значимую единицу (ESU, sensu Moritz, 1994), эндемичную для этой географической области и, насколько известно, фрагментированную на три географических изолята, ограниченных горными вершинами выше 1000 м над уровнем моря. .s.l. в горных массивах Аспромонте, Сила, Поллино. Необходимы дальнейшие исследования для оценки демографической согласованности и моделей генетического разнообразия этих изолятов, а также для лучшего определения наиболее подходящей стратегии управления этой узкой эндемичной линией.
Наконец, наши результаты также могут иметь важное таксономическое значение, которое может иметь решающее значение для сохранения и управления, поскольку приоритеты в стратегиях сохранения определяются на основе статуса и разнообразия видов (см. E.g., Конвенция о международной торговле видами дикой фауны и флоры, находящимися под угрозой исчезновения (СИТЕС), внесена в список видов; Красный список МСОП видов, находящихся под угрозой исчезновения). Отнесение аллопатрических популяций к рангу вида или подвида на основе степени генетической дивергенции может быть проблематичным, поскольку модели репродуктивной изоляции не могут быть оценены в полевых условиях. Тем не менее, учитывая основные теоретические и прикладные последствия, связанные с таксономическим рангом, в этом отношении было предпринято несколько попыток либо путем изучения альтернативных определений концепции вида, либо путем сканирования литературы в поисках правдоподобных пороговых значений генетической дивергенции, чтобы присвоить таксон рангу вида относительно взглядов на млекопитающих см. Baker and Bradley, 2006).По делу D. n. aspromontis , дивергенция последовательности CYTB , которую мы обнаружили относительно ближайшей популяции на северо-востоке Италии, равна или даже превышает расхождения, наблюдаемые у многих сестринских видов млекопитающих и, в частности, грызунов (см., например, Michaux et al., 2002; Baker and Bradley, 2006; Wauters et al., 2017). Кроме того, D. n. aspromontis демонстрирует отчетливые морфологические особенности, касающиеся уникального окраса шерсти (см. выше). Соответственно, популяциям лесной сони на юге Италии действительно можно было присвоить видовой ранг.В этом случае Dryomys aspromontis Von Lehmann, 1964 будет доступным в качестве названия таксона с подходящим общим названием как калабрийская лесная соня, поскольку, насколько известно, ее нынешний ареал в основном ограничен этим регионом. Однако в данном случае необходимо сделать предостережение, основанное, по крайней мере, на одном главном аргументе. Паттерны генетического разнообразия у Dryomys nitedula на уровне всего ареала еще не исследованы. Поскольку существует несколько морфологически определенных единиц (т.е. подвидов), происходящих из географической близости друг к другу в пределах Dryomys nitedula от континентальной Европы до Центральной Азии, тщательное изучение связанных паттернов генетической дивергенции и, что наиболее важно, репродуктивной изоляции может дать сравнительные, но важные знания, чтобы улучшить обоснованные решения о правильном таксономическом ранжировании южно-итальянской линии.
Тем не менее, учитывая основные теоретические и прикладные последствия, связанные с таксономическим рангом, в этом отношении было предпринято несколько попыток либо путем изучения альтернативных определений концепции вида, либо путем сканирования литературы в поисках правдоподобных пороговых значений генетической дивергенции, чтобы присвоить таксон рангу вида относительно взглядов на млекопитающих см. Baker and Bradley, 2006).По делу D. n. aspromontis , дивергенция последовательности CYTB , которую мы обнаружили относительно ближайшей популяции на северо-востоке Италии, равна или даже превышает расхождения, наблюдаемые у многих сестринских видов млекопитающих и, в частности, грызунов (см., например, Michaux et al., 2002; Baker and Bradley, 2006; Wauters et al., 2017). Кроме того, D. n. aspromontis демонстрирует отчетливые морфологические особенности, касающиеся уникального окраса шерсти (см. выше). Соответственно, популяциям лесной сони на юге Италии действительно можно было присвоить видовой ранг.В этом случае Dryomys aspromontis Von Lehmann, 1964 будет доступным в качестве названия таксона с подходящим общим названием как калабрийская лесная соня, поскольку, насколько известно, ее нынешний ареал в основном ограничен этим регионом. Однако в данном случае необходимо сделать предостережение, основанное, по крайней мере, на одном главном аргументе. Паттерны генетического разнообразия у Dryomys nitedula на уровне всего ареала еще не исследованы. Поскольку существует несколько морфологически определенных единиц (т.е. подвидов), происходящих из географической близости друг к другу в пределах Dryomys nitedula от континентальной Европы до Центральной Азии, тщательное изучение связанных паттернов генетической дивергенции и, что наиболее важно, репродуктивной изоляции может дать сравнительные, но важные знания, чтобы улучшить обоснованные решения о правильном таксономическом ранжировании южно-итальянской линии.
Благодарность
Мы благодарим Луку Лапини и А. (Саса) Пеллегрино за любезный обмен фотографиями, а также Марии Кьяре Дефлориан (Музей науки Тренто, Muse), Паоло Паолуччи (Университет Падуи) и Антонио Маццеи (Университет Калабрии) за их помощь в сборе образцов и / или в лаборатории. процедуры. Это исследование было поддержано грантами Министерства образования, университетов и исследований Италии (проект PRIN 2012FRHYRA) и Национального парка Аспромонте.
(Саса) Пеллегрино за любезный обмен фотографиями, а также Марии Кьяре Дефлориан (Музей науки Тренто, Muse), Паоло Паолуччи (Университет Падуи) и Антонио Маццеи (Университет Калабрии) за их помощь в сборе образцов и / или в лаборатории. процедуры. Это исследование было поддержано грантами Министерства образования, университетов и исследований Италии (проект PRIN 2012FRHYRA) и Национального парка Аспромонте.
Обзор воздействия качества лесных местообитаний и нарушения ландшафта на экологию и сохранение видов сони
Адамик П., Крал М. (2008) Утрата гнезд гнездящихся птиц из-за сони (Gliridae, Rodentia).Acta Theriol 53: 185–192. https://doi.org/10.1007/BF03194251
Статья Google ученый
Arnan X, Comas L, Gracia M, Retana J (2014) Состав и использование среды обитания мелких млекопитающих в старовозрастных горных лесах. J Nat Hist 48: 481–494. https://doi.org/10.1080/00222933.2013.800611
Статья Google ученый
Бако Б., Хеккер К. (2006) Факторы, определяющие распространение сосуществующих видов сони (Gliridae, Rodentia).Pol J Ecol 54 (3): 379–386
Google ученый
Bani L, Orioli V, Pisa G, Fagiani S, Dondina O, Fabbri E, Randi E, Sozio G, Mortelliti A (2017) Генетическая структура популяции и расселение лещинной сони с учетом пола ( Muscardinus avellanarius ) ) в непрерывном и фрагментированном ландшафте центральной Италии. Conserv Genet 18: 261–274. https://doi.org/10.1007/s10592-016-0898-2
Статья Google ученый
 org/ScholarlyArticle">
org/ScholarlyArticle">Bani L, Orioli V, Pisa G, Dondina O, Fagiani S, Fabbri E, Randi E, Mortelliti A, Sozio G (2018) Ландшафтные детерминанты генетической дифференциации, инбридинга и генетического дрейфа у ореховой сони ( Muscardinus avellanarius ).Conserv Genet 19: 283–296. https://doi.org/10.1007/s10592-017-0999-6
CAS Статья Google ученый
Берг Л. (1996) Мелкомасштабные изменения в распределении сони Muscardinus avellanarius (Rodentia, Myoxidae) в связи с изменениями растительности. Mammalia 60: 211–216. https://doi.org/10.1515/mamm.1996.60.2.211
Статья Google ученый
Берг Л., Берг Å (1998) Выбор места гнездования сони Muscardinus avellanarius в двух разных ландшафтах.Энн Зул Фен 35: 115–122
Google ученый
Берг Л., Берг Å (1999) Изобилие и выживаемость лещинной сони Muscardinus avellanarius во временной среде обитания кустарников: исследование отлова. Энн Зул Фен 36: 159–165
Google ученый
Bertolino S (2007) Использование микропредприятий садовым сони во время ночной активности. J Zool 272: 176–182
Статья Google ученый
Bertolino S (2017) Распространение и статус убывающей садовой сони Eliomys quercinus .Mam Rev 47: 133–147. https://doi.org/10.1111/mam. 12087
12087
Статья Google ученый
Бертолино С., Кордеро Н., Куррадо I (2003) Домашние хребты и среда обитания садовой сони ( Eliomys quercinus ) в горной среде обитания летом. Acta Zool Acade Sci Hung 49: 11–18
Google ученый
Бибер К. (1995) Распространенное поведение сони съедобной ( Myoxus glis ) на фрагментированном ландшафте в центральной Германии.Гистрикс 6: 257–264. https://doi.org/10.4404/hystrix-6.1-2-4037
Статья Google ученый
Бибер К., Руф Т. (2009) Различия в среде обитания влияют на тактику жизненного цикла импульсного потребителя ресурсов, съедобной сони ( Glis glis ). Поп Эко 51: 481. https://doi.org/10.1007/s10144-009-0140-x
Статья Google ученый
Bright PW (1998) Поведение специализированных видов в коридорах местообитаний: древесный сони избегают пробелов в коридорах.Анимационное поведение 56: 1485–1490. https://doi.org/10.1006/anbe.1998.0921
CAS Статья PubMed PubMed Central Google ученый
Bright PW, Morris PA (1990) Требования к среде обитания сони Muscardinus avellanarius в связи с управлением лесными угодьями в Юго-Западной Англии. Биол Консерв 54: 307–326. https://doi.org/10.1016/0006-3207(90)-D
Статья Google ученый
 org/ScholarlyArticle">
org/ScholarlyArticle">Bright PW, Morris PA (1991) Расположение и гнездовое поведение сони, Muscardinus avellanarius , в разнообразных низкорослых лесах.J Zool 224: 177–190. https://doi.org/10.1111/j.1469-7998.1991.tb04797.x
Статья Google ученый
Bright PW, Morris PA (1992) Расположение и гнездовое поведение сони Muscardinus avellanarius в лесистой местности со стандартами. J Zool 226: 589–600. https://doi.org/10.1111/j.1469-7998.1992.tb07502.x
Статья Google ученый
Bright PW, Morris PA (1993) Кормление сони Muscardinus avellanarius в двух контрастирующих местообитаниях.Дж. Зоол 230: 69–85. https://doi.org/10.1111/J.1469-7998.1993.TB02673.X
Статья Google ученый
Брайт П.В., Моррис П.А. (1994) Перемещение животных в целях сохранения: характеристики сони в зависимости от методов выпуска, происхождения и сезона. J Appl Ecol 31: 699–708. https://doi.org/10.2307/2404160
Статья Google ученый
Bright PW, Morris PA (1996) Почему сони встречаются редко? Пример из природоохранной биологии.Mam Rev 26: 157–187. https://doi.org/10.1111/j.1365-2907.1996.tb00151.x
Статья Google ученый
Bright PW, Mitchell P, Morris PA (1994) Распространение сони: методы обследования, экология островов и выбор участков для сохранения. J Appl Ecol. https://doi.org/10.2307/2404547
J Appl Ecol. https://doi.org/10.2307/2404547
Статья Google ученый
Bright PW, Morris PA, Mitchell P (2006) Справочник по сохранению сони, 2-е изд.English Nature, Питерборо
Google ученый
Büchner S (2008) Распространение обыкновенного соня обыкновенного Muscardinus avellanarius в мозаике местообитаний. Acta Theriol 53: 259–262. https://doi.org/10.1007/BF03193122
Статья Google ученый
Büchner S, Lang J (2014) Die Haselmaus ( Muscardinus avellanarius ) в Германии - Lebensräume, Schutzmaßnahmen und Forschungsbedarf.Säugetierkundliche Informationen 9: 367–377
Статья Google ученый
Capizzi D, Battistini M, Amori G (2002) Анализ лещинной сони, Muscardinus avellanarius , распространение в средиземноморских фрагментированных лесах. Ita J Zool 69: 25–31. https://doi.org/10.1080/11250000209356434
Статья Google ученый
Capizzi D, Battistini M, Amori G (2003) Влияние фрагментации среды обитания и управления лесами на распространение съедобной сони Glis glis .Acta Theriol 48: 359–371. https://doi.org/10.1007/BF03194175
Статья Google ученый
Чанин П. , Губерт Л. (2012) Движение сони обыкновенной ( Muscardinus avellanarius ) в ландшафте, фрагментированном дорогами. Лутра 55: 3–15
, Губерт Л. (2012) Движение сони обыкновенной ( Muscardinus avellanarius ) в ландшафте, фрагментированном дорогами. Лутра 55: 3–15
Google ученый
Cornils JS, Hoelzl F, Rotter B, Bieber C, Ruf T (2017) Сонь съедобный ( Glis glis ) избегайте районов с высокой плотностью их предпочтительного пищевого растения - европейского бука.Front Zool 14:23. https://doi.org/10.1186/s12983-017-0206-0
Статья PubMed PubMed Central Google ученый
Curtis PG, Slay CM, Harris NL, Tyukavina A, Hansen MC (2018) Классификация факторов глобальной потери лесов. Наука 361: 1108–1111. https://doi.org/10.1126/science.aau3445
CAS Статья PubMed Google ученый
Дитц М., Бюхнер С., Хиллен Дж., Шульц Б. (2018) Карта мелких млекопитающих: определение и улучшение крупномасштабной и трансграничной связи местообитаний лещинной сони Muscardinus avellanarius на фрагментированной сельскохозяйственной территории.Biodivers Conserv 27: 1891–1904. https://doi.org/10.1007/s10531-018-1515-0
Статья Google ученый
Дондина О., Катаока Л., Ориоли В., Бани Л. (2016) Как управлять живыми изгородями как эффективными экологическими коридорами для млекопитающих: двухвидовый подход. Сельское хозяйство Ecosyst Environ 231: 283–290. https://doi.org/10.1016/j.agee.2016.07.005
Статья Google ученый
 org/ScholarlyArticle">
org/ScholarlyArticle">Даунс, Северная Каролина, Дин М., Уэллс Д., Воутерс А. (2020) Вытеснение и перемещение лещинного соня ( Muscardinus avellanarius ) в качестве мер по смягчению последствий дорожного строительства.Мам Коммуна 6: 1–9
Google ученый
Фиц Дж., Вайс-Дотц Т. (2012) Оказавшись на острове: последствия фрагментации леса для изменений размеров тела древесного млекопитающего, съедобной сони ( Glis glis ). Pop Ecol 54: 313–320. https://doi.org/10.1007/s10144-012-0310-0
Статья Google ученый
Фиц Дж., Томюк Дж., Лешке В., Вайс-Доц Т., Сегельбахер Г. (2014) Генетические последствия фрагментации леса для высокоспециализированного древесного млекопитающего - съедобной сони.PLoS ONE 9 (2): e88092. https://doi.org/10.1371/journal.pone.0088092
CAS Статья PubMed PubMed Central Google ученый
Figarski T (2009) Przypadek wykorzystywani a nory ziemnej przez koszatkę Dryomys nitedula w trakcie sezonu rozrodczego w Lasach Koneckich. Кулон 14: 140–142
Google ученый
Фигарски Т. (2010) Ochrona popielicy ( Glis glis L., 1766) a sposób zagospodarowania buczyn w Polsce [Консервирование жирной сони ( Glis glis L., 1766) и управление буковыми лесами в Польше]. Chrońmy Przyr Ojcz 66: 39–44
Google ученый
 org/ScholarlyArticle">
org/ScholarlyArticle">Friebe K, Steffens T, Schulz B, Valqui J, Reck H, Hartl G (2018) Значение основных дорог как барьеров и их придорожных местообитаний как потенциальных коридоров для миграции лещинных сони - популяционно-генетическое исследование. Folia Zool 67: 98–109.https://doi.org/10.25225/fozo.v67.i2.a10.2018
Статья Google ученый
Gatter W, Schütt R (2001) Langzeitpopulationsdynamik des Siebenschläfers Myoxus glis в Баден-Вюртемберге - Ein Kleinsäuger als Gewinner der heutigen Waldwirtschaft und des gesellschaftlichen Wandels. Jahreshefte der Gesellschaft für Naturkunde в Вюртемберге 157: 181–210
Google ученый
Global Forest Watch, 2002 г.Глобальная лесная вахта. Институт мировых ресурсов, Вашингтон, округ Колумбия. http://www.globalforestwatch.org. По состоянию на март 2002 г.
Goodwin CE, Hodgson DJ, Al-Fulaij N, Bailey S, Langton S, Mcdonald RA (2017) Схема добровольной регистрации выявляет продолжающееся сокращение популяции лещинных соней в Соединенном Королевстве Muscardinus avellanarius . Mam Rev 47: 183–197. https://doi.org/10.1111/mam.12091
Статья Google ученый
Goodwin CE, Suggitt AJ, Bennie J, Silk MJ, Duffy JP, Al-Fulaij N, Bailey S, Hodgson DJ, McDonald RA (2018) Климат, ландшафт, среда обитания и ассоциации управления лесными угодьями с лещиной соней Muscardinus avellanarius Статус популяции.Mam Rev 48: 209–223. https://doi.org/10.1111/mam.12125
Статья Google ученый
 org/ScholarlyArticle">
org/ScholarlyArticle">Gornung E, Bizzoco D, Colangelo P, Castiglia R (2010) Сравнительное цитогенетическое и генетическое исследование двух итальянских популяций садовой сони Eliomys quercinus L. (Sciuromorpha: Gliridae). Ital J Zool 77: 137–143. https://doi.org/10.1080/112500004107
CAS Статья Google ученый
Grass I, Loos J, Baensch S, Batáry P, Librán-Embid F, Ficiciyan A, Klaus F, Riechers M, Rosa J, Tiede J (2019) Ландшафты связи с разделением / сохранением земли для экосистемных услуг и сохранение биоразнообразия.Люди Nat 1: 262–272. https://doi.org/10.1002/pan3.21
Статья Google ученый
Грегори Р.Д., Воржишек П., Нобл Д.Г., Ван Стриен А., Клванёва А., Итон М., Мейлинг AWG, Джойс А., Фоппен Р.П., Бурфилд И.Дж. (2008) Создание и использование индикаторов популяции птиц в Европе. Bird Conserv Int 18: S223 – S244. https://doi.org/10.1017/S0959270
0312
Статья Google ученый
Gurnell J, Hicks M, Whitbread S (1992) Влияние ухода за порослью на популяции мелких млекопитающих.В кн .: Экология и лесопользование. Springer, pp. 213–232
Hecker K, Bako B, Csorba G (2003) Экология распространения венгерских видов сони на основе национальной системы мониторинга биоразнообразия. Acta Zool Acad Sci Hun 49: 45–54
Google ученый
Herdegen M, Radwan J, Sobczynska U, Dabert M, Konjević D, Schlichter J, Jurczyszyn M (2016) Структура популяции съедобной сони в Польше: роль фрагментации среды обитания и последствия для сохранения.Дж. Зоол 298: 217–224. https://doi.org/10.1111/jzo.12304
Статья Google ученый
Holden-Musser ME, Juškaitis R, Musser GM (2016) Gliridae. В: Wilson DE, Lacher TE, Mittermeier RA (eds) Справочник млекопитающих мира - том 6, зайцеобразные и грызуны I. и книга млекопитающих мира - том 6. Lynx Editions, стр. 838–889
Google ученый
Iannarilli F, Melcore I, Sozio G, Roviani D, Mortelliti A (2017) Модели долгосрочной колонизации и исчезновения лесозависимого грызуна ( Muscardinus avellanarius ) в сильно фрагментированных ландшафтах.Гистрикс. https://doi.org/10.4404/HYSTRIX-28.1-11886
Статья Google ученый
МСОП (2020) Красный список МСОП видов, находящихся под угрозой исчезновения. Версия 2020-1
Iwińska K, Boratyński JS, Tvedi A, Borowski Z (2020) Ежедневное использование насестов съедобной сони в управляемом лесу с преобладанием сосны. Для Ecol Manag 468: 118172
Jurczyszyn M (1997) Wpływ gospodarki leśnej na wyginiecie popielicy, Myoxus glis [L.] w północno-zachodniej Polsce. Badania Fizjograficzne nad Polsk Zachodnią Seria C: Zoologia 44: 71–76
Google ученый
Юрчишин М. (2001) Реинтродукция съедобной сони ( Glis glis ) в ландшафтном парке Сераковски (Польша). Предварительные результаты. Trakya Univ J Sci Res Ser B 2: 111–114
Google ученый
Jurczyszyn M (2006) Использование пространства перемещенными съедобными сонями, Glis glis (L.), в месте их первоначального захвата и месте их выпуска: метод радиотрекинга, примененный в эксперименте по реинтродукции. Pol J Ecol 54: 345–350
Google ученый
Jurczyszyn M (2018) Пищевые и кормовые предпочтения съедобной сони Glis glis на двух участках в Польше. Folia Zool 67: 83–90. https://doi.org/10.25225/fozo.v67.i2.a5.2018
Статья Google ученый
Юшкайтис Р. (1997) Расположение и передвижение обыкновенной сони Muscardinus avellanarius в Литве.Acta Theriol 42: 113–122
Статья Google ученый
Юшкайтис Р. (2005) Влияние высокой плотности гнездовых яиц на популяцию обыкновенной сони Muscardinus avellanarius . Acta Theriol 50: 43–50. https://doi.org/10.1007/BF03192617
Статья Google ученый
Юшкайтис Р. (2006) Взаимодействие между сонями (Gliridae) и птицами, гнездящимися в гнездах.Folia Zool 55: 225–236
Google ученый
Юшкайтис Р. (2007a) Выбор среды обитания в сони обыкновенной Muscardinus avellanarius (L.) в Литве. Baltic For 13: 89–95
Google ученый
Юшкайтис Р. (2007b) Питание сони обыкновенной ( Muscardinus avellanarius ): обзор. Acta Zool Lit 17: 151–159
Статья Google ученый
Юшкайтис Р. (2008) Долгосрочный мониторинг сони обыкновенной: влияние лесопользования на численность.Biodivers Conserv 17: 3559–3565. https://doi.org/10.1007/s10531-008-9470-9
Статья Google ученый
Juškaitis R (2018) Статус сони (Gliridae) в Литве и соседних странах: обзор. Folia Zool 67: 64–68. https://doi.org/10.25225/fozo.v67.i1.a6.2018
Статья Google ученый
Juškaitis R (2020) Орешник лещинный ( Muscardinus avellanarius ) на восстанавливающейся поляне: влияние сплошных рубок и рубок ухода за лесом на долгосрочную динамику численности.Eur J Wildl Res 66:48. https://doi.org/10.1007/s10344-020-01392-z
Статья Google ученый
Юшкайтис Р., Ремейзис Р. (2007a) Летние гнездовья обыкновенной сони Muscardinus avellanarius L. в молодых лесах Литвы. Pol J Ecol 55: 795
Google ученый
Juškaitis R, Remeisis R (2007b) Уловы мышей Micromys minutus и сони обыкновенного Muscardinus avellanarius живут симпатрическими в лесной среде обитания.Acta Theriol 52: 349–354. https://doi.org/10.1007/BF03194232
Статья Google ученый
Юшкайтис Р., Шиожините V (2008) Требования к среде обитания обыкновенной сони ( Muscardinus avellanarius) и толстой сони (Glis glis) в зрелых смешанных лесах Литвы. Ecol Bratisl 27: 143
Google ученый
Юшкайтис Р., Балчяускас Л., Шижините В. (2012) Предпочтение места гнездования лесной сони Dryomys nitedula (Pallas) в северо-западном углу ареала.Pol J Ecol 60: 815–826
Google ученый
Юшкайтис Р., Бальчяускас Л., Шиожините В. (2013) Выбор места гнездования орешниковой соней Muscardinus avellanarius : безопасность важнее еды? Зоол Стад 52:53. https://doi.org/10.1186/1810-522X-52-53
Статья Google ученый
Юшкайтис Р., Балтрунайте Л., Китрите Н. (2016) Кормление в непредсказуемой среде: ежегодные изменения в рационе лещинной сони Muscardinus avellanarius .Mam Res 61: 367–372. https://doi.org/10.1007/s13364-016-0280-2
Статья Google ученый
Karantanis N-E, Rychlik L, Herrel A, Youlatos D (2017) Сравнение древесных походок Muscardinus avellanarius и Glis glis (Gliridae, Rodentia): первый количественный анализ. Mam Study 42: 161–172. https://doi.org/10.3106/041.042.0306
Статья Google ученый
Кекель М., Бюхнер С., Ансорге Х (2012) Зависит ли появление орешниковой сони Muscardinus avellanarius в Восточной Саксонии (Германия) от изолированности и размера среды обитания? Печиана 8: 57–60
Google ученый
Kelm J, Lange A, Schulz B, Göttsche M, Steffens T, Reck H (2015) Как часто лесные млекопитающие добровольно переходят дорогу? Новые сведения о поведении орешниковой сони в придорожных местообитаниях.J Verteb Biol 64: 342–348. https://doi.org/10.25225/fozo.v64.i4.a9.2015
Статья Google ученый
Krofel M, Huber D, Kos I (2011) Рацион евразийской рыси Lynx lynx в северных Динарских горах (Словения и Хорватия). Acta Theriol 56: 315–322. https://doi.org/10.1007/s13364-011-0032-2
Статья Google ученый
Kryŝtufek B, Vohralik V (1994) Распространение лесной сони Dryomys nitedula (Pallas, 1779) (Rodentia, Myoxidae) в Европе.Мам Rev 24: 161–177. https://doi.org/10.1111/j.1365-2907.1994.tb00140.x
Статья Google ученый
Ланг Дж., Бюхнер С., Элерс С., Шульц Б. (2013) Kompensationsmaßnahmen für Haselmäuse im Wald. AFZ Der Wald 10: 14–17
Google ученый
Lebl K, Rotter B, Kürbisch K, Bieber C, Ruf T (2011) Факторы местной окружающей среды влияют на репродуктивные инвестиции самок съедобного соня.J Mam 92: 926–933. https://doi.org/10.1644/10-MAMM-A-225.1
Статья Google ученый
Магомедов М. (2015) Оценка качества среды обитания лесной сони ( Dryomys nitedula ) в Дагестане, Россия: роль кормов и структура растительности. Folia Zool 64: 356–360. https://doi.org/10.25225/fozo.v64.i4.a11.2015
Статья Google ученый
Мянд Р., Тилгар В., Лыхмус А., Лейвиц А. (2005) Обеспечение гнездовых ящиков птицам, гнездящимся в норках - имеет ли значение среда обитания? Biodivers Conserv 14: 1823–1840.https://doi.org/10.1007/s10531-004-1039-7
Статья Google ученый
Марчези Л., Серджио Ф., Педрини П. (2002) Затраты и выгоды от разведения орлиной совы в измененных человеком ландшафтах Bubo bubo . Ibis 144: E164 – E177. https://doi.org/10.1046/j.1474-919X.2002.t01-2-00094_2.x
Статья Google ученый
Марчези Л., Серджио Ф., Педрини П. (2006) Влияние временных изменений в динамике леса на плотность, выбор места для гнезд, рацион и продуктивность неясыти Strix aluco в Альпах.Исследование птиц 53: 310–318. https://doi.org/10.1080/00063650609461447
Статья Google ученый
Марто М., Сара М. (2015) Среда обитания съедобной сони, Glis glis italicus : значение для управления древесными млекопитающими в средиземноморских лесах. Folia Zool 64: 136–150. https://doi.org/10.25225/fozo.v64.i2.a7.2015
Статья Google ученый
Michaux J, Hürner H, Krystufek B, Sarà M, Ribas A, Cornuet JM, Ruch T., Vekhnik V, Renaud S (2019) Генетическая структура европейского лесного вида, съедобной сони ( Glis glis ) : последствия прошлой антропогенной фрагментации лесов? Биол Дж. Линн Соц. 126: 836–851.https://doi.org/10.1093/biolinnean/bly176
Статья Google ученый
Микусиньски Г., Громадски М., Хиларецкий П. (2001) Дятлы как индикаторы разнообразия лесных птиц. Conserv Biol 15: 208–217. https://doi.org/10.1046/j.1523-1739.2001.99236.x
Статья Google ученый
Милаццо А., Фаллетта В., Сара М. (2003) Выбор места обитания жирной сони ( Glis glis italicus ) в лиственных лесах Сицилии.Acta Zool Acad Sci Hung 49: 117–124
Google ученый
Минато С., Ивабучи М., Айба Х, Отаке К., Моррис П. (2012 г.) Помогая сони (японский язык) перейти дорогу. Орикс 46: 325–326. https://doi.org/10.1017/S0030605312000762
Статья Google ученый
Mitchell-Jones AJ, White I (2009) Использование повторных интродукций для восстановления утраченного ареала обитания сони, Muscardinus avellanarius , в Англии.Folia Zool 58: 341–348
Google ученый
Mitchell-Jones AJ, Amori G, Bogdanowicz W, Krystufek B, Reijnders PJH, Spitzenberger F, Stubbe M, Thissen JBM, Vohralik V, Zima J (1999) Атлас европейских млекопитающих, том 3. Academic Press, Лондон
Google ученый
Moher D, Liberati A, Tetzlaff J, Altman DG, Group P (2009) Предпочтительные элементы отчетности для систематических обзоров и метаанализов: заявление PRISMA.PLoS Med 6: e1000097. https://doi.org/10.1371/journal.pmed.1000097
Статья PubMed PubMed Central Google ученый
Морено С., Руко С. (2013) Ответ сообщества мелких млекопитающих на управление средой обитания посредством контролируемого сжигания в охраняемой зоне Средиземного моря. Acta Oecol 49: 1–4. https://doi.org/10.1016/j.actao.2013.02.001
Статья Google ученый
Mortelliti A, Sanzo GS, Boitani L (2009) Суррогатное материнство видов для планирования сохранения: предостережения от сравнения реакции трех древесных грызунов на потерю и фрагментацию среды обитания.Biodivers Conserv 18: 1131–1145. https://doi.org/10.1007/s10531-008-9477-2
Статья Google ученый
Mortelliti A, Amori G, Capizzi D, Cervone C, Fagiani S, Pollini B, Boitani L (2011) Независимые эффекты утраты среды обитания, фрагментации среды обитания и структурной взаимосвязи на распространение двух древесных грызунов. J Appl Ecol 48: 153–162. https://doi.org/10.1111/j.1365-2664.2010.01918.x
Статья Google ученый
Mortelliti A, Santarelli L, Sozio G, Fagiani S, Boitani L (2013) Дальние полевые переходы лещинного соня ( Muscardinus avellanarius ) на фрагментированных ландшафтах.Мам Биол 78: 309–312. https://doi.org/10.1016/j.mambio.2012.09.006
Статья Google ученый
Mortelliti A, Sozio G, Driscoll DA, Bani L, Boitani L, Lindenmayer DB (2014) Популяционные и индивидуальные реакции на размер пятна, изоляцию и качество у лещинной сони. Экосфера 5: 1–21. https://doi.org/10.1890/ES14-00115.1
Статья Google ученый
Моска М., Муха А., Вежбицкий Х (2018) Генетическая дифференциация съедобной сони ( Glis glis ) в польских Судетах: текущий статус исчезающего вида.J Zool 305: 203–211. https://doi.org/10.1111/jzo.12552
Статья Google ученый
Naim DM, Telfer S, Tatman S, Bird S, Kemp SJ, Hughes R, Watts PC (2012) Паттерны генетической дивергенции среди популяций обыкновенной сони, Muscardinus avellanarius в Великобритании. Mol Biol Rep 39: 1205–1215. https://doi.org/10.1007/s11033-011-0850-y
CAS Статья PubMed Google ученый
Naim DM, Telfer S, Tatman S, Bird S, Kemp SJ, Watts PC (2014) Модели передвижения и генетическое разнообразие дикого и реинтродуцированного соня обыкновенного, Muscardinus avellanarius .Genet Mol Res 13: 167–181. https://doi.org/10.4238/2014.January.10.8
CAS Статья PubMed Google ученый
Nakamura-Kojo Y, Kojo N, Ootsuka T., Minami M, Tamate HB (2014) Влияние ресурсов дерева на использование гнездового ящика японской соней Glirulus japonicus . Mam Study 39: 17–26. https://doi.org/10.3106/041.039.0104
Статья Google ученый
Наварро Л.М., Перейра Х.М. (2015) На пути к европейской политике в области повторного вайлдинга.Rewilding Eur Landsc 205
Недялков Н., Попгеоргиев Г., Станева А. (2018) Обновленное распределение неуловимой мышинохвостой сони плотвы, Myomimus roachi Bate, 1937 (Mammalia: Rodentia: Gliridae) в Болгарии. Hist Nat Bulg 29: 3–8
Google ученый
Obuch J, Danko Š, Mihók J, Karaska D, Šimák L (2013) Диета уральской совы ( Strix uralensis ) в Словакии. Raptor J 7: 59–71.https://doi.org/10.2478/srj-2013-0003
Статья Google ученый
Орлов В.Н., Кривоногов Д.М., Черепанова Е.В., Сазанова О.Г., Дмитриев А.И. (2013) Экстенсивный спад распространения сони садовой ( Eliomys quercinus ) и вероятность утраты внутривидового таксономического разнообразия. Зоол Журнал 92: 1349–1358. https://doi.org/10.7868/S004451341311010X
Статья Google ученый
Panchetti F, Sorace A, Amori G, Carpaneto GM (2007) Предпочтение места гнездования обыкновенной сони (M uscardinus avellanarius ) в двух разных типах местообитаний в центральной Италии.Ital J Zool 74: 363–369. https://doi.org/10.1080/11250000701588224
Статья Google ученый
Perry RW, Jenkins JM, Thill RE, Thompson FR III (2018) Долгосрочное воздействие различных методов лесовосстановления на взрослых лесных птиц. Для Ecol Manag 408: 183–194. https://doi.org/10.1016/j.foreco.2011.05.002
Статья Google ученый
Пешев С, Пешев Д, Попов В (2004) Фауна Болгарии, т. 27.Mammalia, BAS, София ( [на болгарском языке с резюме на английском] )
Google ученый
Пилатс В., Пилате Д., Орниканс А., Карклиньш А. (2012) Использование микроместа обитания лесным сони ( Dryomys nitedula ) в бореонеморальном лесу - предварительные результаты. Печяна 8: 77–85
Google ученый
Рамакерс Дж.Дж., Доренбош М., Фоппен Р.П. (2014) Выживание на грани: анализ среды обитания, ориентированный на сохранение, и манипуляции с краями леса для лещинной сони в Нидерландах.Eur J Wildl Res 60: 927–931. https://doi.org/10.1007/s10344-014-0849-5
Статья Google ученый
Руф Т., Фиц Дж., Шлунд В., Бибер С. (2006) Высокая выживаемость в плохие годы: тактика жизненного цикла адаптирована к посеву мачты в съедобной сони. Экология 87: 372–381. https://doi.org/10.1890/05-0672
CAS Статья PubMed Google ученый
Sarà M, Milazzo A, Falletta W, Bellia E (2005) Соревнование по эксплуатации между гнездящимися норками ( Muscardinus avellanarius , Mammalia и Parus caeruleus, Aves) в средиземноморских лесах.J Zool 265: 347–357. https://doi.org/10.1017/S0952836
645X
Статья Google ученый
Scaravelli D, Bassi S (1994) Myoxus glis как пещерное животное. Гистрикс 6: 283–285. https://doi.org/10.4404/hystrix-6.1-2-4041
Статья Google ученый
Schickmann S, Urban A, Kräutler K, Nopp-Mayr U, Hackländer K (2012) Взаимосвязь микофагов мелких млекопитающих и эктомикоризных грибов в первобытных, нарушенных и управляемых горных лесах Центральной Европы.Oecologia 170: 395–409. https://doi.org/10.1007/s00442-012-2303-2
Статья PubMed PubMed Central Google ученый
Ściński M, Borowski Z (2006) Домашние пастбища, места гнездования и динамика численности лесной сони Dryomys nitedula pallas в дубовом грабовом лесу: исследование с отловом и радиоактивным отслеживанием. Pol J Ecol 54: 391–396
Google ученый
Shibata F, Kawamichi T, Nishibayashi K (2004) Ежедневный выбор мест отдыха и их использование японскими сонями.J Mam 85: 30–37. https://doi.org/10.1644/BPR-007
Статья Google ученый
Sozio G, Iannarilli F, Melcore I, Boschetti M, Fipaldini D, Luciani M, Roviani D, Schiavano A, Mortelliti A (2016) Управление лесным хозяйством влияет на индивидуальные и популяционные параметры лещинной сони Muscardinus avellanarius . Мам Биол 81: 96–103. https://doi.org/10.1016/j.mambio.2014.12.006
Статья Google ученый
Trout RC, Brooks SE, Rudlin P, Neil J (2012) Влияние восстановления хвойных плантаций на древнем лесном массиве (PAWS) в Великобритании на среду обитания и местное население ореховой сони ( Muscardinus avellanarius) ).Eur J Wildl Res 58: 635–643. https://doi.org/10.1007/s10344-012-0611-9
Статья Google ученый
Trout RC, Brooks S, Combe FJ, Rudlin P (2018) Различные эффекты периодических экспериментальных схем удаления деревьев на демографию популяции лещинного сони ( Muscardinus avellanarius ) на хвойных плантациях. Folia Zool 67: 110–119. https://doi.org/10.25225/fozo.v67.i2.a7.2018
Статья Google ученый
Tscharntke T, Clough Y, Wanger TC, Jackson L, Motzke I, Perfecto I, Vandermeer J, Whitbread A (2012) Глобальная продовольственная безопасность, сохранение биоразнообразия и будущее интенсификации сельского хозяйства.Биол Консерв 151: 53–59. https://doi.org/10.1016/j.biocon.2012.01.068
Статья Google ученый
Вехник В.А. (2019) Влияние кормовой доступности на воспроизводство у пищевого соня ( Glis glis L., 1766) на восточной периферии ареала. Mam Res 64: 423–434. https://doi.org/10.1007/s13364-019-00425-6
Статья Google ученый
Verbeylen G (2006) Статус и сохранение обыкновенной сони ( Muscardinus avellanarius ) в провинции Лимбург (Фландрия, Бельгия).Лутра 49: 75–88
Google ученый
White IC, Hughes SA (2019) Испытание моста для воссоединения фрагментированной древесной среды обитания лещинной сони Muscardinus avellanarius в заповеднике Бридлсфорд, остров Уайт. UK Conserv Evid 16: 6–11
Google ученый
Уильямс Р.Л., Гуденаф А.Е., Харт А.Г., Стаффорд Р. (2013) Использование долгосрочных записей добровольцев для изучения выбора гнездовых ящиков для сони ( Muscardinus avellanarius ).PLoS ONE 8 (6): e67986. https://doi.org/10.1371/journal.pone.0067986
CAS Статья PubMed PubMed Central Google ученый
Wolton R (2009) Орешниковая соня Muscardinus avellanarius (L.) Выбор места гнездования в живой изгороди. Mammalia 73: 7–12. https://doi.org/10.1515/MAMM.2009.001
Статья Google ученый
Юлатос Д., Карантанис Н.Е., Байрон С.Д., Панютина А. (2015) Захват педали у древесного грызуна связан с поведением над ветвями на тонких субстратах.Дж. Зоол 296: 239–248. https://doi.org/10.1111/jzo.12237
Статья Google ученый
Zapponi L, Del Bianco M, Luiselli L, Catorci A, Bologna MA (2013) Оценка воздействия экологических требований на чувствительность к фрагментации леса у двух древесных грызунов.